Kinetics of Glycoxidation of Bovine Serum Albumin by Glucose, Fructose and Ribose and Its Prevention by Food Components

Abstract
:1. Introduction
2. Results
2.1. Kinetics of Glycoxidation
2.2. Protection against Glycoxidation




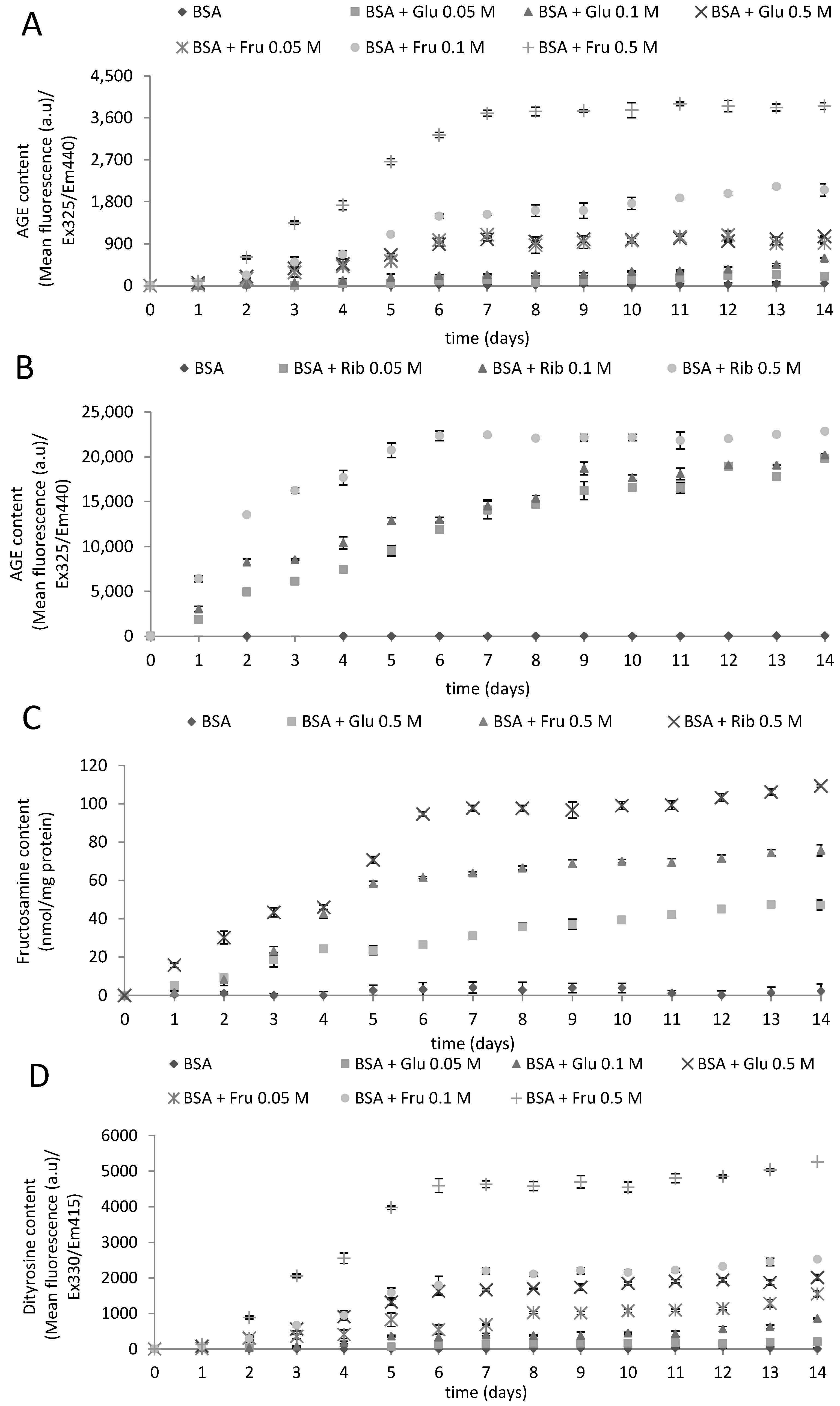{kind=link}
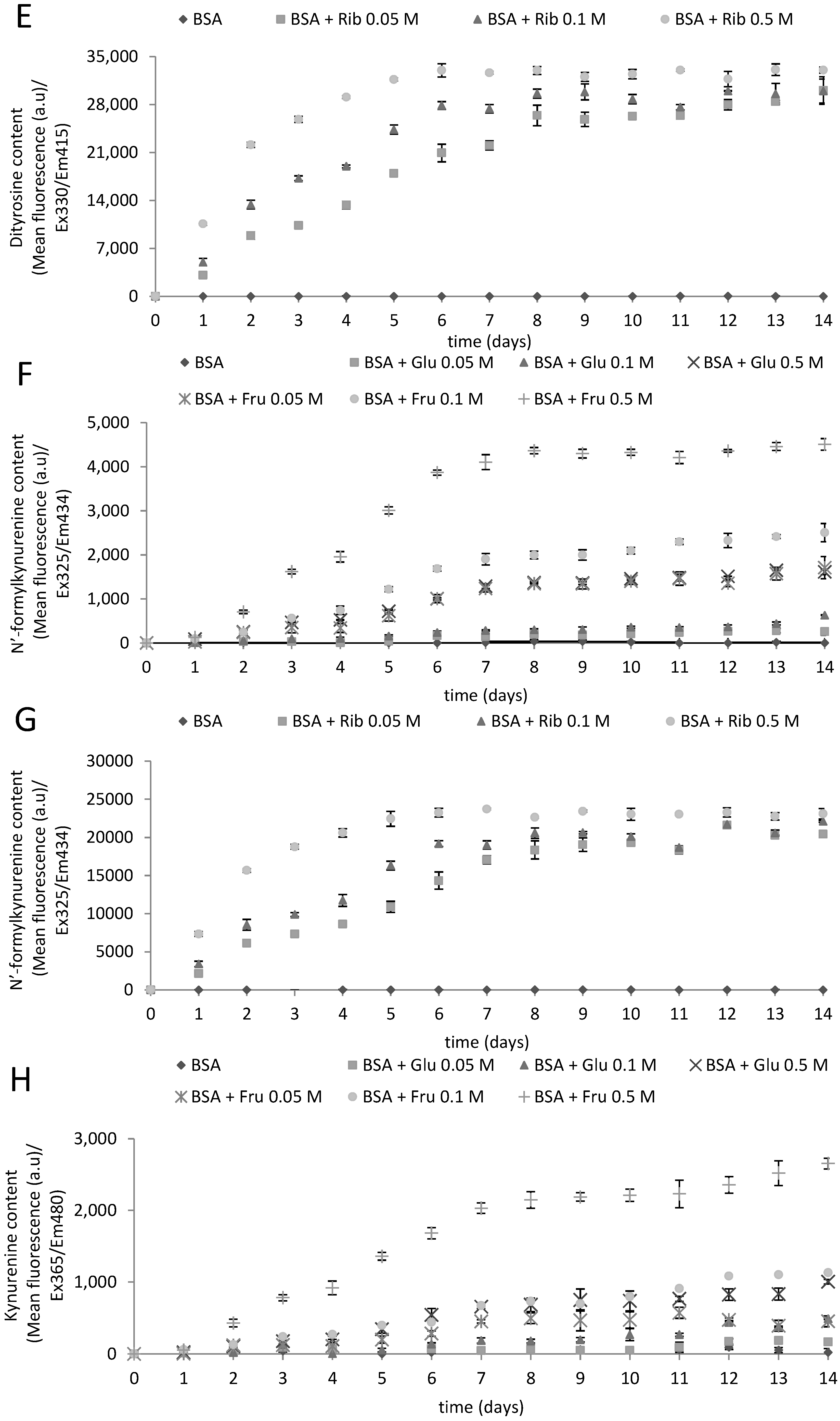{kind=link}
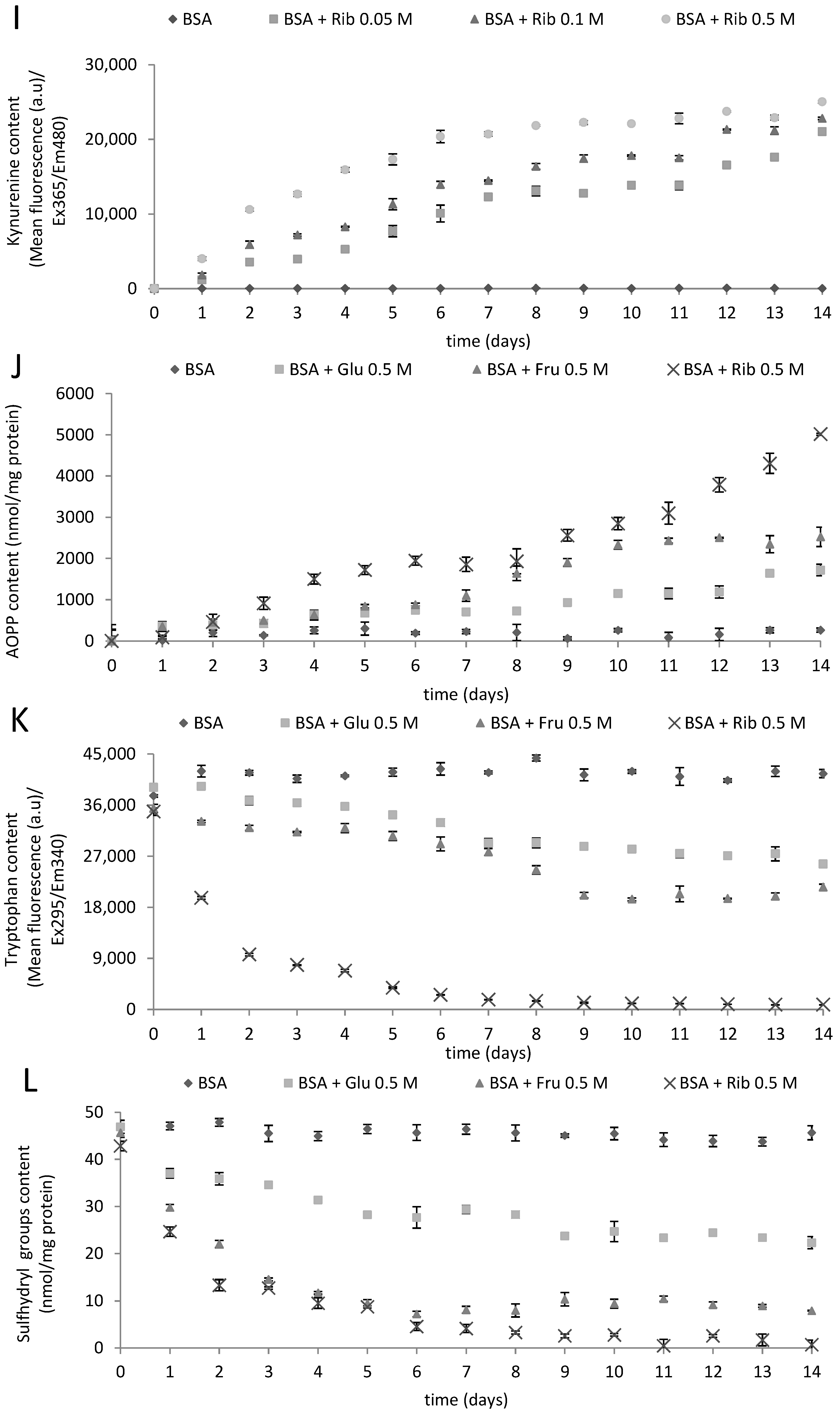{kind=link}
| Additive | AGE | Dityrosine | N'-formylkynurenine | Kynurenine | AOPP |
|---|---|---|---|---|---|
| BSA, no sugar | 3.55 ± 5.16 * | 3.46 ± 5.61 * | 1.57 ± 5.37 * | 7.27 ± 10.06 * | 17.25 ± 6.58 # |
| BSA + glucose, no additive | 100 | 100 | 100 | 100 | 100 |
| DMSO | 103.78 ± 10.32 | 100.56 ± 10.59 | 103.45 ± 10.91 | 232.47 ± 23.09 # | 91.11 ± 9.34 |
| Standard Antiglycating Agents | |||||
| Aminoguanidine | 41.91 ± 2.69 * | 29.39 ± 4.13 * | 38.07 ± 3.07 * | 66.58 ± 10.87 Δ | 41.31 ± 4.87 # |
| Metformin | 112.75 ± 3.34 Δ | 116.00 ± 2.79 # | 124.59 ± 3.01 # | 119.83 ± 6.21 Δ | 118.21 ± 9.02 |
| Pyridoxine | <0 * | <0 * | <0 * | 193.42 ± 14.04 # | 6.19 ± 4.27 * |
| Antioxidants | |||||
| Captopril | 34.52 ± 7.55 # | 31.85 ± 6.81 # | 37.62 ± 7.22 # | 87.27 ± 16.74 | 33.66 ± 6.14 # |
| Tiron | 31.3 ± 10.73 # | 28.91 ± 10.92 # | 33.12 ± 11.23 # | 72.55 ± 15.79 Δ | 65.87 ± 13.88 |
| Metal Chelators | |||||
| EDTA | 98.14 ± 2.27 | 92.47 ± 3.40 | 99.53 ± 4.81 | 87.01 ± 7.95 | 106.87 ± 13.37 |
| DETAPA | 135.79 ± 2.61 # | 127.51 ± 1.63 Δ | 125.27 ± 4.11 Δ | 94.94 ± 2.24 | 107.18 ± 17.16 |
| NTA | 124.59 ± 9.51 Δ | 113.80 ± 8.24 | 112.30 ± 8.92 | 94.03 ± 13.82 | 109.73 ± 8.69 |
| Amino Acids and Derivatives, Peptides | |||||
| Arginine | 120.14 ± 9.56 | 106.81 ± 9.16 | 108.58 ± 8.69 | 104.33 ± 14.37 | 107.86 ± 14.18 |
| Carnosine | 122.00 ± 6.13 Δ | 120.20 ± 6.46 Δ | 135.06 ± 7.29 # | 152.47 ± 9.75 # | 127.47 ± 31.73 |
| Cysteamine | 20.19 ± 9.35 # | 19.16 ± 9.82 # | 22.82 ± 10.75 # | 48.48 ± 22.52 Δ | 46.15 ± 10.14 Δ |
| Glutathione oxidized | 136.72 ± 10.56 Δ | 140.81 ± 11.13 Δ | 144.95 ± 11.84 Δ | 149.44 ± 20.76 | 110.25 ± 8.37 |
| Glutathione reduced | 51.49 ± 13.28 Δ | 49.26 ± 10.97 # | 49.61 ± 10.87 # | 111.08 ± 25.92 | 83.66 ± 8.70 |
| Glycine | 140.89 ± 20.65 | 143.49 ± 18.41 | 148.09 ± 21.45 | 148.40 ± 28.52 | 110.04 ± 10.32 |
| Organic Acids | |||||
| 1-Cyano-4-hydroxycinnamic acid | 4.89 ± 1.60 * | 4.59 ± 1.26 * | 6.12 ± 2.29 * | 19.48 ± 5.36 * | 31.62 ± 9.55 # |
| 4-Hydroxy cinnamic acid | 94.70 ± 7.05 | 97.77 ± 6.09 | 104.29 ± 6.23 | 92.12 ± 12.43 | 89.07 ± 9.50 |
| Lipoic acid | 89.23 ± 2.41 Δ | 89.35 ± 2.42 # | 97.02 ± 3.17 | 97.73 ± 2.80 | 91.71 ± 9.89 |
| Para-aminobenzoic acid | 152.04 ± 18.58 Δ | 150.09 ± 16.01 Δ | 163.83 ± 17.49 Δ | 149.39 ± 15.80 Δ | 129.74 ± 25.42 |
| Pyruvic acid | 82.29 ± 3.11 # | 84.56 ± 2.68 # | 91.26 ± 2.88 Δ | 73.94 ± 6.16 # | 94.54 ± 2.27 |
| Quinic acid | 72.93 ± 8.44 Δ | 75.19 ± 8.59 Δ | 79.49 ± 8.34 Δ | 56.54 ± 12.28 Δ | 88.53 ± 11.99 |
| Organic Polybases | |||||
| Spermidine | 143.88 ± 4.11 # | 137.43 ± 3.23 # | 151.7 ± 3.39 * | 233.07 ± 8.83 * | 117.47 ± 16.58 |
| Spermine | 144.22 ± 19.98 | 139.62 ± 23.29 | 153.53 ± 22.79 | 195.06 ± 35.58 Δ | 118.37 ± 13.14 |
| Polyphenols | |||||
| Caffeic acid | <0 * | <0 * | <0 * | 180.71 ± 9.74 # | 43.38 ± 14.08 Δ |
| Ellagic acid | 96.47 ± 6.16 | 58.82 ± 4.37 # | 92.01 ± 6.56 | 107.71 ± 35.95 | 103.99 ± 41.12 |
| Ferulic acid | <0 * | <0 * | 29.34 ± 7.35 # | 156.28 ± 19.39 Δ | 53.27 ± 9.22 Δ |
| Gallic acid | 460.03 ± 9.73 * | 249.02 ± 14.12 # | 432.07 ± 11.04 * | 586.37 ± 49.71 # | 615.09 ± 24.62 * |
| Genistein | 27.46 ± 0.97 * | 23.86 ± 0.79 * | 27.77 ± 1.43 * | 46.85 ± 1.77 * | 44.43 ± 10.14 Δ |
| Kaempferol | 20.77 ± 2.09 * | 13.98 ± 1.58 * | 20.18 ± 2.43 * | 22.12 ± 4.38 * | 45.97 ± 18.44 Δ |
| Naringin | 28.90 ± 0.44 * | 22.05 ± 0.17 * | 24.38 ± 0.35 * | 103.39 ± 4.68 | 35.87 ± 11.61 Δ |
| Propyl gallate | <0 * | <0 * | <0 * | 198.18 ± 5.78 * | 71.84 ± 20.08 |
| Quercitrin | 3.37 ± 0.38 * | 1.11 ± 0.28 * | 1.01 ± 0.17 * | 3.31 ± 0.58 * | 47.35 ± 14.16 Δ |
| Rutin | 2.94 ± 0.35 * | 1.15 ± 0.34 * | 1.26 ± 0.23 * | 2.79 ± 0.14 * | 42.26 ± 5.76 # |
| Additive | AGE | Dityrosine | N'-formylkynurenine | Kynurenine | AOPP |
|---|---|---|---|---|---|
| BSA, no sugar | 0.67 ± 0.97 * | 0.65 ± 1.05 * | 0.29 ± 0.99 * | 1.52 ± 2.10 * | 8.91 ± 3.40 * |
| BSA + fructose, no additive | 100 | 100 | 100 | 100 | 100 |
| DMSO | 68.54 ± 4.64 # | 62.86 ± 3.89 # | 67.52 ± 4.32 # | 134.00 ± 10.30 Δ | 70.55 ± 3.61 # |
| Standard Antiglycating Agents | |||||
| Aminoguanidine | 19.53 ± 1.58 * | 17.76 ± 0.667 * | 19.20 ± 0.75 * | 39.31 ± 1.96 * | 44.15 ± 3.33 # |
| Metformin | 82.45 ± 5.02 Δ | 83.67 ± 5.14 Δ | 87.36 ± 4.91 Δ | 75.52 ± 7.14 Δ | 123.16 ± 13.28 |
| Pyridoxine | <0 * | <0 * | <0 * | 181.88 ± 3.75 * | 34.26 ± 4.68 # |
| Antioxidants | |||||
| Captopril | 27.87 ± 0.78 * | 25.15 ± 0.78 * | 28.50 ± 0.93 * | 59.26 ± 2.32 * | 16.28 ± 3.17 * |
| Tiron | 97.32 ± 0.70 Δ | 104.07 ± 0.78 # | 103.62 ± 0.98 Δ | 105.68 ± 1.72 Δ | 44.98 ± 4.78 # |
| Metal Chelators | |||||
| EDTA | 98.57 ± 2.99 | 97.09 ± 2.99 | 99.2 ± 1.50 | 99.76 ± 2.01 | 111.12 ± 12.93 |
| DETAPA | 63.07 ± 3.04 # | 59.98 ± 2.92 # | 58.47 ± 2.79 # | 49.35 ± 7.83 # | 87.21 ± 4.25 Δ |
| NTA | 107.64 ± 3.03 Δ | 102.55 ± 3.12 | 101.09 ± 2.80 | 93.19 ± 4.03 Δ | 109.27 ± 5.26 |
| Amino Acids and Derivatives, Peptides | |||||
| Arginine | 93.26 ± 3.56 Δ | 87.62 ± 3.70 Δ | 89.68 ± 4.06 Δ | 81.59 ± 4.85 Δ | 107.2 ± 41.66 |
| Carnosine | 97.12 ± 5.10 | 96.28 ± 4.74 | 103.05 ± 5.13 | 104.80 ± 8.01 | 109.53 ± 18.10 |
| Cysteamine | 54.44 ± 1.83 * | 53.36 ± 1.78 * | 56.81 ± 2.18 * | 71.95 ± 4.41 # | 46.10 ± 13.99 Δ |
| Glutathione oxidized | 111.61 ± 1.64 # | 113.09 ± 2.61 # | 115.42 ± 2.35 # | 114.77 ± 3.19 # | 120.90 ± 16.07 |
| Glutathione reduced | 78.10 ± 7.50 Δ | 78.39 ± 6.58 Δ | 77.68 ± 6.80 Δ | 69.38 ± 8.54 Δ | 129.12 ± 2.89 # |
| Glycine | 118.53 ± 7.88 Δ | 118.7 ± 7.03 Δ | 120.96 ± 6.82 Δ | 120.01 ± 8.89 Δ | 102.15 ± 21.96 |
| Organic Acids | |||||
| 1-Cyano-4-hydroxycinnamic acid | 5.57 ± 0.16 * | 5.14 ± 0.13 * | 6.27 ± 0.16 * | 20.81 ± 0.59 * | 52.02 ± 13.55 Δ |
| 4-Hydroxycinnamic acid | 88.85 ± 3.17 Δ | 90.26 ± 3.39 Δ | 94.86 ± 3.67 | 93.19 ± 5.23 | 63.30 ± 17.30 |
| Lipoic acid | 132.99 ± 4.67 # | 130.7 ± 4.22 # | 137.82 ± 4.29 # | 129.87 ± 4.37 # | 150.17 ± 8.03 # |
| Para-aminobenzoic acid | 102.75 ± 4.31 | 97.82 ± 3.74 | 106.23 ± 4.13 | 107.31 ± 4.52 | 111.79 ± 9.52 |
| Pyruvic acid | 61.70 ± 7.64 # | 62.90 ± 7.79 # | 65.43 ± 8.03 # | 39.36 ± 9.49 # | 87.62 ± 5.22 |
| Quinic acid | 96.86 ± 2.00 | 98.96 ± 1.73 | 104.02 ± 1.80 Δ | 98.33 ± 2.61 | 85.14 ± 6.43 |
| Organic Polybases | |||||
| Spermidine | 101.10 ± 1.64 | 94.96 ± 3.30 | 102.77 ± 2.56 | 153.21 ± 8.47 # | 93.39 ± 12.04 |
| Spermine | 120.74 ± 7.01 Δ | 118.59 ± 7.42 Δ | 125.58 ± 7.11 Δ | 150.61 ± 5.94 # | 98.82 ± 49.59 |
| Polyphenols | |||||
| Caffeic acid | <0 * | <0 * | <0 * | 239.70 ± 68.36 Δ | 79.04 ± 6.87 Δ |
| Ellagic acid | 31.62 ± 4.73 * | 21.03 ± 3.68 * | 29.80 ± 4.41 * | 60.44 ± 7.81 # | 129.61 ± 5.19 Δ |
| Ferulic acid | <0 * | <0 * | 0.42 ± 3.25 * | 92.74 ± 5.87 | 43.52 ± 3.32 # |
| Gallic acid | 59.54 ± 1.65 * | 33.05 ± 1.20 * | 55.43 ± 1.18 * | 217.27 ± 2.94 * | 92.99 ± 21.89 |
| Genistein | 14.88 ± 0.61 * | 12.64 ± 0.34 * | 14.97 ± 0.49 * | 20.47 ± 0.25 * | 69.88 ± 10.88 Δ |
| Kaempferol | 14.36 ± 1.15 * | 9.98 ± 0.77 * | 13.63 ± 1.10 * | 18.55 ± 2.11 * | 64.21 ± 7.72 Δ |
| Naringin | 27.03 ± 0.30 * | 21.53 ± 0.38 * | 24.90 ± 0.97 * | 99.74 ± 3.01 | 76.34 ± 4.21 Δ |
| Propyl gallate | <0 * | <0 * | <0 * | 54.36 ± 6.62 # | 41.08 ± 11.75 Δ |
| Quercitrin | 2.62 ± 0.26 * | 1.77 ± 0.22 * | 2.08 ± 0.30 * | 1.84 ± 0.23 * | 43.63 ± 2.57 * |
| Rutin | 2.48 ± 0.11 * | 1.82 ± 0.07 * | 2.20 ± 0.10 * | 1.82 ± 0.09 * | 50.28 ± 7.56 # |
| Additive | AGE | Dityrosine | N'-formylkynurenine | Kynurenine | AOPP |
|---|---|---|---|---|---|
| BSA, no sugar | 0.67 ± 0.97 * | 0.65 ± 1.05 * | 0.29 ± 0.99 * | 1.52 ± 2.10 * | 1.82 ± 0.70 * |
| BSA + ribose, no additive | 100 | 100 | 100 | 100 | 100 |
| DMSO | 92.02 ± 0.64 # | 92.38 ± 0.67 # | 93.92 ± 0.84 # | 113.33 ± 0.96 * | 109.68 ± 0.87 # |
| Standard Antiglycating Agents | |||||
| Aminoguanidine | 78.07 ± 1.53 * | 70.13 ± 0.32 * | 79.71 ± 0.44 * | 80.21 ± 2.06 * | 64.35 ± 4.39 # |
| Metformin | 99.66 ± 1.77 | 100.52 ± 1.67 | 105.57 ± 1.63 Δ | 91.19 ± 3.96 | 80.84 ± 4.72 Δ |
| Pyridoxine | <0 * | <0 * | <0 * | 95.09 ± 2.42 | 10.02 ± 3.45 * |
| Antioxidants | |||||
| Captopril | 93.66 ± 0.41 * | 94.57 ± 0.80 # | 98.39 ± 0.69 Δ | 102.39 ± 2.97 | 91.08 ± 2.02 Δ |
| Tiron | 97.32 ± 0.70 Δ | 104.07 ± 0.78 # | 103.62 ± 0.98 Δ | 105.68 ± 1.72 Δ | 51.85 ± 4.39 # |
| Metal Chelators | |||||
| EDTA | 98.67 ± 0.59 | 97.64 ± 6.72 | 99.29 ± 7.77 | 100.31 ± 8.44 | 96.71 ± 0.74 |
| DETAPA | 104.09 ± 0.48 # | 99.73 ± 0.62 | 100.59 ± 0.78 | 978.65 ± 1.79 | 76.42 ± 3.41 # |
| NTA | 111.73 ± 0.79 * | 107.94 ± 1.07 # | 107.87 ± 0.80 # | 104.63 ± 2.52 | 86.01 ± 2.59 Δ |
| Amino Acids and Derivatives, Peptides | |||||
| Arginine | 89.83 ± 0.86 # | 89.06 ± 0.80 * | 87.61 ± 0.96 * | 97.67 ± 1.91 | 86.78 ± 4.06 Δ |
| Carnosine | 93.56 ± 0.43 * | 93.68 ± 0.35 * | 99.28 ± 0.24 Δ | 102.90 ± 0.77 | 77.96 ± 3.48 # |
| Cysteamine | 78.55 ± 1.87 # | 79.47 ± 1.85 # | 82.13 ± 1.98 # | 104.22 ± 3.52 | 93.21 ± 1.68 Δ |
| Glutathione oxidized | 83.63 ± 4.11 # | 91.42 ± 3.07 Δ | 87.73 ± 4.38 Δ | 106.09 ± 4.06 | 95.55 ± 3.30 |
| Glutathione reduced | 85.58 ± 1.45 # | 91.99 ± 1.15 # | 86.17 ± 1.16 # | 103.70 ± 3.42 | 109.26 ± 3.92 |
| Glycine | 99.46 ± 0.55 | 103.24 ± 0.24 * | 104.19 ± 0.99 # | 109.24 ± 2.23 # | 89.25 ± 3.25 Δ |
| Organic Acids | |||||
| 1-Cyano-4-hydroxycinnamic acid | 18.29 ± 0.11 * | 16.46 ± 0.04 * | 18.54 ± 0.05 * | 35.31 ± 0.19 * | 33.38 ± 5.90 # |
| 4-Hydroxycinnamic acid | 102.44 ± 0.28 # | 104.63 ± 0.40 # | 110.93 ± 0.26 * | 97.48 ± 0.27 # | 96.52 ± 3.52 |
| Lipoic acid | 105.45 ± 0.61 # | 102.81 ± 0.83 Δ | 108.73 ± 0.35 * | 111.00 ± 2.01 # | 83.08 ± 5.73 Δ |
| Para-aminobenzoic acid | 85.53 ± 1.05 * | 80.87 ± 1.05 * | 87.46 ± 0.93 * | 99.43 ± 1.78 | 82.13 ± 3.72 Δ |
| Pyruvate | 104.70 ± 2.98 | 105.50 ± 3.08 | 111.00 ± 3.13 Δ | 99.18 ± 4.47 | 98.48 ± 8.42 |
| Quinic acid | 108.40 ± 0.90 # | 110.98 ± 0.90 # | 116.92 ± 1.11 * | 107.96 ± 0.80 # | 105.43 ± 3.09 |
| Organic Polybases | |||||
| Spermidine | 38.84 ± 0.37 * | 36.07 ± 0.21 * | 40.85 ± 0.39 * | 87.18 ± 0.65 * | 44.69 ± 5.23 # |
| Spermine | 32.06 ± 1.86 * | 29.54 ± 1.80 * | 33.71 ± 1.87 * | 71.84 ± 0.59 * | 67.50 ± 3.97 # |
| Polyphenols | |||||
| Caffeic acid | 45.63 ± 22.36 Δ | 56.37 ± 24.57 Δ | 48.62 ± 12.47 # | 96.67 ± 6.67 | 64.26 ± 0.88 * |
| Ellagic acid | 74.18 ± 2.54 # | 74.19 ± 2.95 # | 74.74 ± 2.85 # | 44.32 ± 1.84 * | 77.97 ± 6.66 Δ |
| Ferulic acid | 14.04 ± 0.52 * | 28.05 ± 0.49 * | 18.08 ± 0.51 * | 99.22 ± 1.05 | 41.60 ± 2.21 * |
| Gallic acid | 73.48 ± 1.87 * | 86.01 ± 1.52 # | 79.09 ± 1.49 * | 64.21 ± 1.38 * | 81.57 ± 3.22 Δ |
| Genistein | 29.40 ± 0.99 * | 28.48 ± 1.02 * | 30.75 ± 0.93 * | 33.91 ± 0.82 * | 17.42 ± 3.80 * |
| Kaempferol | 29.23 ± 1.95 * | 24.84 ± 1.96 * | 29.68 ± 1.88 * | 19.98 ± 1.81 * | 84.23 ± 6.49 |
| Naringin | 53.60 ± 1.21 * | 50.86 ± 1.11 * | 53.28 ± 1.20 * | 68.38 ± 0.70 * | 58.63 ± 4.45 # |
| Propyl gallate | 66.71 ± 1.60 * | 55.39 ± 2.09 * | 65.65 ± 1.77 * | 81.82 ± 1.30 * | 55.23 ± 3.53 # |
| Quercitrin | 14.20 ± 0.15 * | 12.13 ± 0.13 * | 14.19 ± 0.18 * | 8.37 ± 0.07 * | 50.17 ± 3.76 # |
| Rutin | 15.79 ± 0.30 * | 13.40 ± 0.35 * | 15.78 ± 0.35 * | 7.88 ± 0.14 * | 64.22 ± 6.26 Δ |
| Additive | Glucose | Fructose | Ribose |
|---|---|---|---|
| BSA + sugar, no additive | 100 | 100 | 100 |
| DMSO | 101.31 ± 2.37 | 99.90 ± 2.09 | 101.58 ± 1.89 |
| Standard Antiglycating Agents | |||
| Aminoguanidine | 74.41 ± 0.44 * | 70.63 ± 1.683 * | 88.98 ± 1.00 * |
| Metformin | 96.57 ± 0.46 # | 97.74 ± 2.836 | 99.35 ± 0.54 |
| Pyridoxine | 43.35 ± 2.55 * | 43.08 ± 0.16 * | 71.85 ± 1.86 * |
| Antioxidants | |||
| Captopril | 84.93 ± 1.02 * | 86.56 ± 1.46 * | 98.45 ± 1.92 |
| Tiron | 83.10 ± 0.76 * | 94.41 ± 0.86 # | 98.14 ± 1.73 |
| Metal Chelators | |||
| NTA | 95.35 ± 1.61 Δ | 97.78 ± 0.51 | 100.31 ± 0.71 |
| Amino Acids and Derivatives, Peptides | |||
| Arginine | 97.16 ± 5.84 | 98.35 ± 3.39 | 98.79 ± 0.58 Δ |
| Carnosine | 93.19 ± 2.04 # | 92.55 ± 1.84 # | 95.36 ± 1.44 # |
| Cysteamine | 82.25 ± 0.94 * | 86.00 ± 0.84 * | 88.43 ± 1.93 * |
| Glutathione oxidized | 95.69 ± 0.69 # | 99.24 ± 1.27 | 100.73 ± 0.67 |
| Glutathione reduced | 88.75 ± 2.77 # | 93.19 ± 3.52 Δ | 96.71 ± 0.93 Δ |
| Glycine | 95.61 ± 1.16 # | 99.62 ± 1.35 | 97.51 ± 1.22 Δ |
| Organic Acids | |||
| 1-Cyano-4-hydroxycinnamic acid | 66.41 ± 2.25 * | 72.08 ± 1.89 * | 89.86 ± 0.61 * |
| 4-Hydroxycinnamic acid | 100.64 ± 3.46 | 97.73 ± 0.98 | 101.17 ± 3.11 |
| Lipoic acid | 100.51 ± 8.54 | 96.19 ± 1.23 Δ | 100.78 ± 3.55 |
| Para-aminobenzoic acid | 102.88 ± 2.92 | 93.71 ± 4.11 | 105.51 ± 0.58 * |
| Pyruvic acid | 84.34 ± 1.06 * | 87.07 ± 0.27 * | 98.94 ± 0.50 Δ |
| Quinic acid | 97.05 ± 3.35 | 99.05 ± 2.67 | 98.35 ± 1.87 |
| Organic Polybases | |||
| Spermidine | 103.5 ± 0.65 Δ | 96.48 ± 2.82 | 92.4 ± 0.20 * |
| Polyphenols | |||
| Caffeic acid | 46.8 ± 1.39 * | 61.52 ± 1.31 * | 86.04 ± 0.42 * |
| Ellagic acid | 94.53 ± 0.31 # | 89.76 ± 2.97 # | 100.05 ± 0.55 |
| Ferulic acid | 56.83 ± 3.53 * | 60.96 ± 0.87 * | 85.12 ± 2.72 * |
| Gallic acid | 115.89 ± 4.67 # | 94.30 ± 1.88 Δ | 95.66 ± 1.55 # |
| Genistein | 61.43 ± 2.95 * | 66.39 ± 0.23 * | 86.23 ± 1.10 * |
| Naringin | 57.83 ± 4.10 * | 69.61 ± 1.03 * | 92.52 ± 0.42 * |
| Propyl gallate | 57.38 ± 1.53 * | 63.13 ± 1.83 * | 83.97 ± 1.15 * |
| Rutin | 62.11 ± 3.98 * | 67.99 ± 0.64 * | 77.25 ± 0.61 * |
| Additive | Glucose | Fructose | Ribose |
|---|---|---|---|
| BSA + sugar, no additive | 100 | 100 | 100 |
| DMSO | 98.68 ± 0.76 Δ | 99.18 ± 2.79 | 99.03 ± 0.87 |
| Standard Antiglycating Agents | |||
| Aminoguanidine | 75.38 ± 0.31 * | 79.34 ± 0.85 * | 79.1 ± 4.69 # |
| Metformin | 100.01 ± 2.20 | 98.58 ± 0.32 Δ | 99.39 ± 2.77 |
| Pyridoxine | 53.24 ± 0.69 * | 72.82 ± 1.18 * | 74.48 ± 2.69 * |
| Antioxidants | |||
| Captopril | 88.08 ± 0.06 * | 89.35 ± 2.01 * | 98.06 ± 5.92 |
| Tiron | 94.79 ± 0.48 * | 95.00 ± 2.73 Δ | 95.44 ± 2.47 Δ |
| Metal Chelators | |||
| NTA | 101.66 ± 0.10 * | 100.86 ± 0.69 | 105.87 ± 2.26 |
| Amino Acids and Derivatives, Peptides | |||
| Arginine | 100.65 ± 2.44 | 97.34 ± 2.49 | 95.61 ± 0.72 # |
| Carnosine | 101.20 ± 3.82 | 87.54 ± 0.41 * | 92.33 ± 4.76 |
| Cysteamine | 93.64 ± 1.32 # | 94.56 ± 1.41 # | 92.99 ± 8.31 |
| Glutathione oxidized | 101.24 ± 0.43 Δ | 99.79 ± 2.77 | 102.68 ± 1.00 Δ |
| Glutathione reduced | 98.65 ± 2.79 | 96.47 ± 1.39 Δ | 91.23 ± 3.57 Δ |
| Glycine | 97.25 ± 0.01 * | 96.67 ± 1.15 Δ | 98.33 ± 3.85 Δ |
| Organic Acids | |||
| 1-Cyano-4-hydroxycinnamic acid | 86.38 ± 0.27 * | 82.21 ± 0.46 * | 82.88 ± 2.30 * |
| 4-Hydroxycinnamic acid | 102.02 ± 2.58 | 102.53 ± 1.71 | 97.06 ± 5.98 |
| Lipoic acid | 98.40 ± 0.12 * | 97.9 ± 3.49 | 98.98 ± 0.04 |
| Para-aminobenzoic acid | 104.78 ± 1.51 # | 97.53 ± 1.20 Δ | 102.57 ± 0.59 Δ |
| Pyruvic acid | 98.20 ± 1.71 | 91.35 ± 0.87 * | 102.62 ± 0.69 Δ |
| Quinic acid | 97.65 ± 0.63 # | 98.50 ± 2.12 | 97.62 ± 2.64 |
| Organic Polybases | |||
| Spermidine | 99.73 ± 1.04 | 97.32 ± 3.51 | 93.11 ± 0.53 # |
| Polyphenols | |||
| Caffeic acid | 50.43 ± 0.19 * | 66.40 ± 1.82 * | 79.48 ± 0.99 * |
| Ellagic acid | 101.48 ± 0.472 # | 94.17 ± 2.664 Δ | 101.90 ± 0.74 |
| Ferulic acid | 59.00 ± 0.16 * | 62.31 ± 2.63 * | 81.26 ± 1.52 * |
| Gallic acid | 116.07 ± 0.13 * | 100.52 ± 0.46 | 92.50 ± 4.35 Δ |
| Genistein | 77.97 ± 1.69 * | 86.8 ± 0.28 * | 84.08 ± 1.92 * |
| Naringin | 78.20 ± 0.06 * | 72.53 ± 2.96 * | 91.43 ± 0.11 * |
| Propyl gallate | 53.15 ± 0.20 * | 65.76 ± 1.01 * | 73.83 ± 1.42 * |
| Rutin | 69.32 ± 0.30 * | 70.28 ± 0.18 * | 79.61 ± 4.06 # |
| Parameters of Glycoxidation | AGEs (E) | AGEs (F) | Dityrosine (F) | N'-formylkynurenine (F) | Kynurenine (F) | Protein Carbonyls (F) |
|---|---|---|---|---|---|---|
| Glucose | ||||||
| AGEs (F) | 0.926 * | |||||
| Dityrosine (F) | 0.963 * | 0.962 * | ||||
| N'-formylkynurenine (F) | 0.450 | 0.598 | 0.636 | |||
| Kynurenine (F) | 0.000 | −0.037 | −0.074 | 0.090 | ||
| AOPP (S) | 0.107 | −0.037 | 0.074 | 0.378 | 0.750 | |
| Carbonyl protein (F) | 0.857 ∆ | 0.815 ∆ | 0.852 ∆ | 0.504 | −0.143 | |
| Glycated albumin (E) | 0.857 ∆ | 0.963 * | 0.926 # | 0.738 | −0.071 | 0.821 ∆ |
| Fructose | ||||||
| AGEs (F) | 0.926 # | |||||
| Dityrosine (F) | 0.926 # | 1.000 * | ||||
| N'-formylkynurenine (F) | 0.901 # | 0.972 * | 0.972 * | |||
| Kynurenine (F) | −0.107 | 0.074 | 0.074 | 0.036 | ||
| AOPP (S) | 0.393 | 0.630 | 0.630 | 0.559 | 0.679 | |
| Carbonyl protein (F) | 0.964 * | 0.926 # | 0.926 # | 0.847 ∆ | 0.000 | |
| Glycated albumin (E) | 0.786 ∆ | 0.926 # | 0.926 # | 0.847 ∆ | 0.074 | 0.857 ∆ |
| Ribose | ||||||
| AGEs (F) | 0.786 ∆ | |||||
| Dityrosine (F) | 0.536 | 0.857 ∆ | ||||
| N'-formylkynurenine (F) | 0.714 | 0.964 * | 0.893 # | |||
| Kynurenine (F) | −0.143 | −0.071 | 0.250 | 0.143 | ||
| AOPP (S) | 0.679 | 0.500 | 0.571 | 0.393 | −0.107 | |
| Carbonyl protein (F) | 0.357 | 0.536 | 0.607 | 0.607 | −0.036 | |
| Glycated albumin (E) | 0.321 | 0.214 | 0.143 | 0.250 | −0.321 | 0.821 ∆ |
2.3. Validity of Fluorometric and Spectrophotometric Measures of Glycoxidation
3. Discussion
4. Experimental Section
4.1. Reagents
4.2. Glycation Conditions
4.3. Fluorometric Estimates of Glycoxidation
4.4. Amadori Product
4.5. AOPP and Thiol Assay
4.6. ELISA-Based AGEs, Carbonyl Groups and Bovine Glycated Albumin Assays
4.7. Prevention of Glycoxidation
4.8. Statistics
5. Conclusions
Acknowledgments
Author Contributions
Abbreviations
| AGEs | advanced glycation end products |
| AOPP | advanced oxidation protein products |
| BSA | bovine serum albumin |
| EDTA | ethylenediaminetetraacetic acid |
| DETAPA | diethylenetriaminepentaacetic acid |
| DMSO | dimethylsulfoxide |
| NBT | nitroblue tetrazolium |
| NTA | nitrilotriacetic acid |
| ROS | reactive oxygen species |
Conflicts of interest
References
- Vistoli, G.; de Maddis, D.; Cipak, A.; Zarkovic, N.; Carini, M.; Aldini, G. Advanced glycoxidation and lipoxidation end products (AGEs and ALEs): An overview of their mechanisms of formation. Free Radic. Res. 2013, 47 (Suppl. 1), 3–27. [Google Scholar] [CrossRef] [PubMed]
- Peppa, M.; Uribarri, J.; Vlassara, H. Aging and glycoxidant stress. Hormones (Athens) 2008, 7, 123–132. [Google Scholar]
- Rondeau, P.; Bourdon, E. The glycation of albumin; structural and functional impacts. Biochimie 2011, 93, 645–658. [Google Scholar]
- Muthenna, P.; Akileshwari, C.; Saraswat, M.; Bhanuprakash Reddy, G. Inhibition of advanced glycation end-product formation on eye lens protein by rutin. Br. J. Nutr. 2012, 107, 941–949. [Google Scholar] [CrossRef] [PubMed]
- Aldini, G.; Vistoli, G.; Stefek, M.; Chondrogianni, N.; Grune, T.; Sereikaite, J.; Sadowska-Bartosz, I.; Bartosz, G. Molecular strategies to prevent; inhibit; and degrade advanced glycoxidation and advanced lipoxidation end products. Free Radic. Res. 2013, 47 (Suppl. 1), 93–137. [Google Scholar] [PubMed]
- Sadowska-Bartosz, I.; Galiniak, S.; Bartosz, G.; Rachel, M. Oxidative modification of proteins in pediatric cystic fibrosis with bacterial infections. Oxid. Med. Cell Longev. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Castellani, R.; Smith, M.A.; Richey, P.L.; Perry, G. Glycoxidation and oxidative stress in Parkinson disease and diffuse Lewy body disease. Brain Res. 1996, 737, 195–200. [Google Scholar] [CrossRef]
- Rondeau, P.; Navarra, G.; Cacciabaudo, F.; Leone, M.; Bourdon, E.; Militello, V. Thermal aggregation of glycated bovine serum albumin. Biochim. Biophys. Acta. 2010, 1804, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, E.; Boehm, B.O.; Süssmuth, S.D.; Kientsch-Engel, R.; Sperfeld, A.; Ludolph, A.C.; Tumani, H. The advanced glycation end-product N epsilon-(carboxymethyl)lysine level is elevated in cerebrospinal fluid of patients with amyotrophic lateral sclerosis. Neurosci. Lett. 2004, 371, 226–229. [Google Scholar] [CrossRef] [PubMed]
- Arasteh, A.; Farahi, S.; Habibi-Rezaei, M.; Moosavi-Movahedi, A.A. Glycated albumin: An overview of the in vitro models of an in vivo potential disease marker. J. Diabetes Metab. Disord. 2014, 13. [Google Scholar] [CrossRef]
- Ledesma-Osuna, A.I.; Ramos-Clamon, G.; Vazquez-More, L. Characterization of bovine serum albumin glycated with glucose, galactose and lactose. Acta Biochim. Pol. 2008, 55, 491–497. [Google Scholar] [PubMed]
- Kawasaki, T.; Akanuma, H.; Yamanouchi, T. Increased fructose concentrations in blood and urine in patients with diabetes. Diabetes Care 2012, 25, 353–357. [Google Scholar] [CrossRef]
- Seuffer, R. A new method for the determination of sugars in cerebrospinal fluid. Klin. Chem. Klin. Biochem. 1977, 15, 663–668. [Google Scholar]
- Eijnde, B.O.; van Leemputte, M.; Brouns, F.; van der Vusse, G.J.; Labarque, V.; Ramaekers, M.; van Schuylenberg, R.; Verbessem, P.; Wijnen, H.; Hespel, P. No effects of oral ribose supplementation on repeated maximal exercise and de novo ATP resynthesis. J. Appl. Physiol. 2001, 91, 2275–2281. [Google Scholar] [PubMed]
- Salahuddin, P.; Rabbani, G.; Khan, R.H. The role of advanced glycation end products in various types of neurodegenerative disease: A therapeutic approach. Cell. Mol. Biol. Lett. 2014, 19, 407–437. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.; Shao, X.; Chen, H.; Ho, C.T.; Sang, S. Genistein inhibits advanced glycation end product formation by trapping methylglyoxal. Chem. Res. Toxicol. 2011, 24, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Ramkissoon, J.S.; Mahomoodally, M.F.; Ahmed, N.; Subratty, A.H. Antioxidant and anti-glycation activities correlates with phenolic composition of tropical medicinal herbs. Asian Pac. J. Trop. Biomed. 2013, 6, 561–569. [Google Scholar] [CrossRef]
- Vlassopoulos, A.; Lean, M.E.; Combet, E. Protein-phenolic interactions and inhibition of glycation—Combining a systematic review and experimental models for enhanced physiological relevance. Food Funct. 2014, 5, 2646–2655. [Google Scholar] [CrossRef] [PubMed]
- Verzelloni, E.; Pellacani, C.; Tagliazucchi, D.; Tagliaferri, S.; Calani, L.; Costa, L.G.; Brighenti, F.; Borges, G.; Crozier, A.; Conte, A.; et al. Antiglycative and neuroprotective activity of colon-derived polyphenol catabolites. Mol. Nutr. Food Res. 2011, 55 (Suppl. 1), S35–S43. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, S.; Chen, F.; Wang, M. Treatment of proteins with dietary polyphenols lowers the formation of AGEs and AGE-induced toxicity. Food Funct. 2014, 24, 2656–2661. [Google Scholar] [CrossRef]
- Frizzell, N.; Baynes, J.W. Chelation therapy for the management of diabetic complications; a hypothesis and a proposal for clinical laboratory assessment of metal ion homeostasis in plasma. Clin. Chem. Lab. Med. 2014, 52, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Nagai, R.; Murray, D.B.; Metz, T.O.; Baynes, J.W. Chelation: A fundamental mechanism of action of AGE inhibitors, AGE breakers, and other inhibitors of diabetes complications. Diabetes 2012, 61, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Anguizola, J.; Matsuda, R.; Barnaby, O.S.; Hoy, K.S.; Wa, C.; Debolt, E.; Koke, M.; Hage, D.S. Review: Glycation of human serum albumin. Clin. Chim. Acta 2013, 425, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Bourdon, N.; Loreau, N.; Blache, D. Glucose and free radicals impair the antioxidant properties of serum albumin. FASEB J. 1999, 13, 233–244. [Google Scholar] [PubMed]
- Luers, L.; Rysiewski, K.; Dumpitak, C.; Birkmann, E. Kinetics of advanced glycation end products formation on bovine serum albumin with various reducing sugars and dicarbonyl compounds in equimolar ratios. Rejuvenation Res. 2012, 15, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Suárez, G.; Rajaram, R.; Oronsky, A.L.; Gawinowicz, M.A. Nonenzymatic glycation of bovine serum albumin by fructose (fructation). Comparison with the Maillard reaction initiated by glucose. J. Biol. Chem. 1989, 264, 3674–3679. [Google Scholar] [PubMed]
- Han, C.; Lu, Y.; Wei, Y.; Liu, Y.; He, R. d-Ribose induces cellular protein glycation and impairs mouse spatial cognition. PLoS One 2011, 6, e24623. [Google Scholar] [CrossRef]
- Wei, Y.; Chen, L.; Chen, J.; Ge, L.; He, R.Q. Rapid glycation with d-ribose induces globular amyloid-like aggregations of BSA with high cytotoxicity to SH-SY5Y cells. BMC Cell Biol. 2009, 10. [Google Scholar] [CrossRef]
- Tanaka, S.; Avigad, G.; Brodsky, B.; Eikenberry, E.F. Glycation induces expansion of the molecular packing of collagen. J. Mol. Biol. 1988, 203, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Luciano Viviani, G.; Puddu, A.; Sacchi, G.; Garuti, A.; Storace, D.; Durante, A.; Monacelli, F.; Odetti, P. Glycated fetal calf serum affects the viability of an insulin-secreting cell line in vitro. Metabolism 2008, 57, 163–169. [Google Scholar] [CrossRef]
- Vallianou, N.; Evangelopoulos, A.; Koutalas, P. α-Lipoic acid and diabetic neuropathy. Rev. Diab. Stud. 2009, 6, 230–236. [Google Scholar] [CrossRef]
- Tarwadi, K.V.; Agte, V.V. Effect of micronutrients on methylglyoxal-mediated in vitro glycation of albumin. Biol. Trace Elem. Res. 2011, 143, 717–725. [Google Scholar] [CrossRef]
- Bournival, J.; Francoeur, M.A.; Renaud, J.; Martinoli, M.G. Quercetin and sesamin protect neuronal PC12 cells from high-glucose-induced oxidation; nitrosative stress; and apoptosis. Rejuvenation Res. 2012, 15, 322–333. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Chen, X. Structures required of polyphenols for inhibiting advanced glycation end products formation. Curr. Drug Metab. 2013, 14, 414–431. [Google Scholar] [CrossRef] [PubMed]
- Obrenovich, M.E.; Nair, N.G.; Beyaz, A.; Aliev, G.; Reddy, V.P. The role of polyphenolic antioxidants in health, disease, and aging. Rejuvenation Res. 2010, 13, 631–643. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Saha, C. A review on structure-affinity relationship of dietary flavonoids with serum albumins. J. Biomol. Struct. Dyn. 2014, 32, 1132–1147. [Google Scholar] [CrossRef] [PubMed]
- Briski, K.P.; Patil, G.D. Induction of Fos immunoreactivity labeling in rat forebrain metabolic loci by caudal fourth ventricular infusion of the monocarboxylate transporter inhibitor, alpha-cyano-4-hydroxycinnamic acid. Neuroendocrinology 2005, 82, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Sadowska-Bartosz, I.; Adamczyk-Sowa, M.; Galiniak, S.; Mucha, S.; Pierzchala, K.; Bartosz, G. Oxidative modification of serum proteins in multiple sclerosis. Neurochem. Int. 2013, 63, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.; Baker, J. Assay of serum fructosamine: Internal vs. external standardization. Clin. Chem. 1987, 33, 1955–1956. [Google Scholar] [PubMed]
- Mironova, R.; Niwa, T.; Handzhiyski, Y.; Sredovska, A.; Ivanow, I. Evidence for non-enzymatic glycosylation of Escherichia coli chromosomal DNA. Mol. Microbiol. 2005, 55, 1801–1811. [Google Scholar] [CrossRef] [PubMed]
- Witko-Sarsat, V.; Friedlander, M.; Capeillere-Blandin, C.; Nguyen-Khoa, T.; Nguyen, A.T.; Zingraff, J.; Jungers, P.; Descamps-Latscha, B. Advanced oxidation protein products as a novel marker of oxidative stress in uremia. Kidney Int. 1996, 49, 1304–1313. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadowska-Bartosz, I.; Galiniak, S.; Bartosz, G. Kinetics of Glycoxidation of Bovine Serum Albumin by Glucose, Fructose and Ribose and Its Prevention by Food Components. Molecules 2014, 19, 18828-18849. https://doi.org/10.3390/molecules191118828
Sadowska-Bartosz I, Galiniak S, Bartosz G. Kinetics of Glycoxidation of Bovine Serum Albumin by Glucose, Fructose and Ribose and Its Prevention by Food Components. Molecules. 2014; 19(11):18828-18849. https://doi.org/10.3390/molecules191118828
Chicago/Turabian StyleSadowska-Bartosz, Izabela, Sabina Galiniak, and Grzegorz Bartosz. 2014. "Kinetics of Glycoxidation of Bovine Serum Albumin by Glucose, Fructose and Ribose and Its Prevention by Food Components" Molecules 19, no. 11: 18828-18849. https://doi.org/10.3390/molecules191118828