


Abstract
The hemoglobin vesicle (HbV) is an artificial oxygen carrier that encapsulates a concentrated Hb solution in lipid vesicles (liposomes). The pharmacokinetic properties of HbV were investigated in mice and rats. With use of HbV in which the internal Hb was labeled with 125I (125I-HbV) and cell-free 125I-Hb, it was found that encapsulation of Hb increased the half-life by 30 times, accompanied by decreased distribution in both the liver and kidney. The half-life of HbV was increased, and the uptake clearance for the liver and spleen were decreased with increasing doses of HbV. In an in vitro study, the specific uptake and degradation of HbV in RAW 264.7 cells were found, but this was not the case for parenchymal and endothelial cells. The pharmacokinetics of HbV components (internal Hb and liposomal lipid) were also investigated using 125I-HbV and 3H-HbV (liposomal cholesterol was radiolabeled with tritium-3). The time courses for the plasma concentration curves of 125I-HbV, 3H-HbV, and iron derived from HbV suggest that HbV maintain an intact structure in the blood circulation up to 24 h after injection. 125I-HbV and 3H-HbV were distributed mainly to the liver and spleen. Internal Hb disappeared from both the liver and spleen 5 days after injection, and the liposomal cholesterol disappeared at approximately 14 days. Internal Hb was excreted into the urine and cholesterol into feces via biliary excretion. These results suggest that the HbV has a reasonable blood retention and metabolic and excretion performance and could be used as an oxygen carrier.
Blood transfusions are absolutely essential for resuscitation from massive bleeding after a surgical procedure. However, there is the potential of mismatching, and they can introduce certain infectious diseases such as hepatitis, human immunodeficiency virus, or West Nile virus, which are threats despite the development of the nucleic acid amplification test. In addition, donated red blood cells (RBCs) for blood transfusions can only be stored for a period of 3 weeks in Japan. To overcome these problems, blood substitutes would be highly desirable and have been under development worldwide (Keipert, 1995; Winslow, 2005).
The hemoglobin vesicle (HbV) is an artificial cellular hemoglobin-based oxygen carrier with polyethylene glycol (PEG), in which phospholipid vesicles encapsulating highly concentrated human Hb serve as oxygen carriers with oxygen transport characteristics that are comparable to those of RBCs. In fact, the pharmacological effects of HbV, injected into rats with hemorrhagic shock, have been reported to be equivalent to those of RBCs (Sakai et al., 2004b, 2009). In addition, HbV have been shown to possess a number of positive characteristics such as the absence of viral contamination (Sakai et al., 1993; Abe et al., 2006), a long-term storage period of more than 2 years at room temperature (Goda et al., 1998; Sakai et al., 2000b; Abe et al., 2007), and low toxicity [blood compatibility, and suppressed nephrotoxicity induced by the dimeric form of Hb and hypertension induced by the direct interaction of Hb with nitric oxide (NO) and CO] (Sakai et al., 2000a). Moreover, HbV suspended in a solution of human serum albumin can be used to regulate rheological properties (e.g., viscosity and colloid osmotic pressure) (Sakai et al., 2000c, 2004b). On the basis of these facts, the use of HbV is predicted to be superior to that of a conventional blood transfusion.
Preclinical pharmacokinetic studies of HbV are essential to evaluate the safety and efficacy of HbV. To sustain the pharmacological effect of HbV as artificial oxygen carriers, prolonged oxygen delivery is a required property. In fact, the plasma retention time of free Hb, when isolated from RBCs, is surprisingly short (half-life of ∼0.5–1.5 h). In clinical situations, the total infused dose of RBCs given to patients can be considerable (e.g., for hemorrhagic shock or transfusion during an operation). Because the lipid content of HbV is more than 100 times higher than that of other liposomal preparations such as AmBisome or Doxil, massive amounts of Hb and lipid components can be infused when HbV are substituted for RBCs.
Free Hb molecules can trigger numerous side effects, such as renal toxicity, hypertension, and tissue damage induced by the Fenton reaction, which is mediated by heme (iron) (Balla et al., 2005). On the other hand, high levels of lipid components in the bloodstream, especially cholesterol, are risk factors for kidney disease, arterial sclerosis, and hyperlipidemia (Gröne and Gröne, 2008). Despite the large body of pharmacological evidence for the HbV as an artificial oxygen carrier, little is known concerning its pharmacokinetic properties, especially the fate of each component after injection. As of this writing, metabolism studies of HbV components have only been done by histopathological examination and blood serum biochemistry (Sakai et al., 2001, 2009). However, it is difficult to distinguish exogenous lipid components, derived from HbV, from endogenous substances. Biodistribution of HbV has been examined by 99mTc-labeled HbV (Sou et al., 2005); however, no pharmacokinetic studies of Hb and lipids from administration to excretion were performed.
In the present study, we report on an evaluation of the pharmacokinetic properties of the HbV and its components, from the standpoint of stability in the blood circulation and the metabolism and excretion of each component, in support of its use as an oxygen carrier. For this purpose, we used two different radiolabeled HbV, 125I-HbV in which the enclosed Hb was radiolabeled and 3H-HbV in which the lipid component (cholesterol) was radiolabeled.
Materials and Methods
Materials. An Hb solution was purified from outdated donated blood that was provided by the Japanese Red Cross Society (Tokyo, Japan). Pyridoxal 5′-phosphate was purchased from Sigma-Aldrich (St. Louis, MO). 1,2-Dipalmitoyl-sn-glycero-3-phosphatidylcholine (DPPC), cholesterol, and 1,5-bis-O-hexadecyl-N-succinyl-l-glutamate were purchased from Nippon Fine Chemical Co. Ltd. (Osaka, Japan). 1,2-Distearoyl-sn-glycero-3-phosphatidyl-ethanolamine-N-PEG was purchased from NOF Corporation (Tokyo, Japan). Recombinant human serum albumin (rHSA) was given by NIPRO (Osaka, Japan). Williams' E medium (WE medium) and RPMI 1640 medium were purchased from Sigma-Aldrich. Dulbecco's modified Eagle's medium and penicillin-streptomycin were purchased from Invitrogen (Carlsbad, CA).
Preparation of HbV. HbV were prepared under sterile conditions as reported previously (Sakai et al., 1997). The resulting encapsulated Hb (38 g/dl) contained 14.7 mM pyridoxal 5′-phosphate as an allosteric effector to regulate P50 to 25 to 28 Torr. The lipid bilayer comprised a mixture of DPPC, cholesterol, and 1,5-bis-O-hexadecyl-N-succinyl-l-glutamate at a molar ratio of 5:5:1 and 1,2-distearoyl-sn-glycero-3-phosphatidyl-ethanolamine-N-PEG (0.3 mol%). The averaged particle diameter was 250 to 280 nm. The HbV were suspended in a physiological salt solution at [Hb] (10 g/dl) and [lipids] (6–7 g/dl), filter-sterilized (450 nm pore size, Dismic; Toyo-Roshi, Tokyo, Japan), and purged with N2 before storage.
In Vivo Experiment.Animals. All animal experiments were undertaken in accordance with the guideline principles and procedures of Kumamoto University for the care and use of laboratory animals. Experiments were performed with male ddY mice (28–30 g b.wt.; Japan SLC, Inc., Shizuoka, Japan) and male Sprague-Dawley rats (180–210 g b.wt.; Kyudou Co., Kumamoto, Japan). All animals were maintained under conventional housing conditions, with food and water ad libitum in a temperature-controlled room with a 12-h dark/light cycle. Before the pharmacokinetic studies using 125I-Hb or 125I-HbV, all of the animals were given water containing 5 mM sodium iodide (NaI) to avoid specific accumulation of the isotope in the glandula thyreoidea until the end of the experiment.
Preparation of radiolabeled Hb and HbV.125I-Hb or 125I-HbV were prepared by incubating 200 μl of Hb or HbV (100 mM Hb) with 125-I as Na125I (10 μl) (GE Healthcare, Little Chalfont, Buckinghamshire, UK) in an Iodo-Gen (1,3,4,6-tetrachoro-3α,6α-diphenylglycoluril) tube for 30 min at room temperature. 125I-Hb and 125I-HbV were then isolated from free 125I by passage through a PD-10 column (Pfizer, Inc., Uppsala, Sweden). More than 97% of the total iodine was bound to internal Hb in HbV. 3H-HbV were prepared by mixing HbV (1 ml) with [1,2-3H]cholesterol solution (40 μl) (PerkinElmer Life and Analytical Sciences, Yokohama, Japan) and incubated for 12 h at room temperature. 3H-HbV were filtered through a sterilized filter to remove aggregates (pore size, 450 nm). The incubation of 3H-HbV in serum (24 h, 37°C) revealed that 3H failed to completely dissociate from the HbV. Before use in pharmacokinetic experiments, all of the samples were mixed with unlabeled protein (Hb or HbV) to adjust the target Hb concentration. In addition, rHSA was added to correspond to the colloid osmotic pressure (Sakai et al., 1997).
Administration and collection of blood and organs in mice. ddY mice received a single injection of 125I-Hb (1 mg of Hb/kg), 125I-HbV (1, 10, 200, and 1400 mg of Hb/kg), or 3H-HbV (1400 mg of Hb/kg) containing 5% rHSA in the tail vein under ether anesthesia. At each time after the injection of labeled protein, blood was collected from the inferior vena cava, and plasma was obtained by centrifugation (3000g, 5 min). After collection of blood, the animal was sacrificed for excision of organs. Urine and feces were collected at fixed intervals in a metabolic cage.
Administration and collection of blood and organs in rats. All of the Sprague-Dawley rats were anesthetized with pentobarbital and received a single injection of 125I-HbV (10, 200, and 1400 mg of Hb/kg) or 3H-HbV (1400 mg of Hb/kg) containing 5% rHSA. The blood was collected from the tail vein, and plasma was obtained by centrifugation (3000g, 5 min). At each time after an injection of 125I-HbV or 3H-HbV, the three animals were sacrificed for collection of organs, which were rinsed with saline. Urine and feces were collected at fixed intervals in a metabolic cage.
Measurement of 125I radioactivity. To remove degraded protein and free 125I, 1% bovine serum albumin (BSA) and 40% trichloroacetic acid (TCA) were added to the plasma, and pellets were obtained by centrifugation (1000g, 10 min). The organs, urine, and feces were weighed on an electronic balance. 125I radioactivity was counted using a liquid scintillation counter (ARC-5000; Aloka, Tokyo, Japan).
Measurement of 3H radioactivity. The plasma samples were ultracentrifuged to collect intact HbV (50,000g, 30 min) (Sakai et al., 2001). Collected HbV were solubilized in a mixture of Soluene 350 (PerkinElmer Life and Analytical Sciences) and isopropyl alcohol (at a ratio of 1:1) for 24 h at 50°C and decolorized by treatment with H2O2. The organ samples were rinsed with saline, minced, and solubilized in Soluene 350 for 24 h at 50°C. Urine and feces were also weighed and solubilized in Soluene 350. Radioactivity was determined using a liquid scintillation counter (LSC-5121; Aloka) with Hionic-Fluor (PerkinElmer Life and Analytical Sciences).
Measurement of iron concentration. The iron concentration was calculated from atomic emission spectrometric analysis (Ubest-35; Jasco, Tokyo, Japan) using an absorbance of 415 nm. The specific iron concentration derived from HbV was calculated by subtraction of the plasma iron concentration without HbV injection from that with HbV injection. At the same time, we confirmed that the plasma iron level hardly changed without HbV in mice during the experimental periods.
In Vitro Experiment.Separation of hepatic cells. Separation of hepatic cells from ddY mice was performed using a method similar to that described in a previous report (Nakajou et al., 2005). The portal vein was cannulated with a polyethylene catheter, and the liver was perfused with Gey's balanced salt solution buffer, pH 7.5, without Ca2+ for 10 min at flow rate of 5 ml/min and then with Gey's balanced salt solution buffer, pH 7.5, containing 125 units/ml collagenase and Ca2+ for 3 min at 37°C. The liver was suspended in ice-cold 1% BSA. The cell suspension was centrifuged at 20g for 2 min, and the precipitate and supernatant were collected. Parenchymal and endothelial cells were isolated from the precipitate and supernatant as follows.
Isolation and culture of hepatic parenchymal cells. The precipitate was resuspended in 1% BSA and again centrifuged at 20g for 2 min. The cells in the pellet were resuspended in 10 ml of phosphate-buffered saline (PBS) and layered on top of a two-step Percoll gradient. The gradient consisted of 25% (v/v) Percoll (top) and 50% (v/v) Percoll (bottom). The gradient was centrifuged at 3000g for 30 min, and the intermediate zone was collected. The enriched hepatic parenchymal cells were suspended in WE medium (FCS+) supplemented with 0.1 mg/ml streptomycin and 100 IU/ml penicillin. Aliquots of the cell suspension were seeded into each well (5 × 105 per well) of collagenase-coated 24-multiwell plates. The multiwell plates were incubated for 60 min at 37°C in a CO2 incubator. Each well was washed three times with 1 ml of PBS to remove nonadherent cells, and then further incubated for 12 h with 1 ml of WE medium (FCS+), which was replaced by 1 ml of fresh WE medium (FCS–), followed by a 4-h incubation before the experiment.
Isolation and Culture of Hepatic Endothelial Cells. The supernatant was suspended in 10 ml of RPMI 1640 medium (FCS–) and cultured on a plastic plate for 20 min (37°C) to remove Kupffer cells. The supernatant in the plastic plate was then seeded into each well (2 × 106 per well) of fibronectin-coated 24-multiwell plates. The multiwell plates were incubated for 60 min at 37°C in a CO2 incubator. Each well was washed twice with 1 ml of PBS to remove nonadherent cells and parenchymal cells and then further incubated for 2 h with 1 ml of RPMI 1640 medium (FCS+), which was replaced by 1 ml of fresh RPMI 1640 medium (FCS–), followed by a 4-h incubation before the experiment.
Culture of RAW 264.7 cells. RAW 264.7 cells suspended in Dulbecco's modified Eagle's medium (FCS+) were seeded into each well (5 × 105 per well) of 24-multiwell plates. The multiwell plates were incubated for 24 h at 37°C in a CO2 incubator. Each well was washed twice with 1 ml of PBS, which was replaced by 1 ml of fresh Dulbecco's modified Eagle's medium (FCS–), followed by a 4-h incubation before the experiment.
Cell assays. The parenchymal, endothelial, and RAW 264.7 cells in each well were incubated with various concentrations of 125I-HbV for examination, with or without an excess of corresponding unlabeled HbV. After a 6-h incubation, 0.575 ml of culture medium was removed from each well and mixed with 0.15 ml of 40% TCA and 0.15 ml of 0.7 M AgNO3 in a vortex mixer, followed by centrifugation (1000g, 10 min). The resulting supernatant (0.25 ml) was used to determine TCA-soluble radioactivity, which was taken as an index of the extent of cellular degradation. The remaining cells in each well were washed five times with 1 ml of PBS. The cells were lysed at 37°C for 30 min with 1 ml of 0.1 M NaOH. One portion was used to determine the radioactivity as the cell-associated ligand; the other was used to determine the cellular protein content.
Data analysis. Pharmacokinetic analyses after HbV or Hb administration was performed based on a two-compartment model. Pharmacokinetic parameters were calculated by fitting using MULTI, a normal least-squares program (Yamaoka et al., 1981). The uptake clearance (CLuptake) was calculated, as described in a previous report using integration plot analysis at designated times (from 1 to 30 min) during which the efflux and/or elimination of radioactivity from tissues were negligible (Murata et al., 1998). Data are shown as mean ± S.D. for the indicated number of animals. The overall differences between groups were determined by one-way analysis of variance. A value of p < 0.05 was considered to indicate statistical significance.
Results
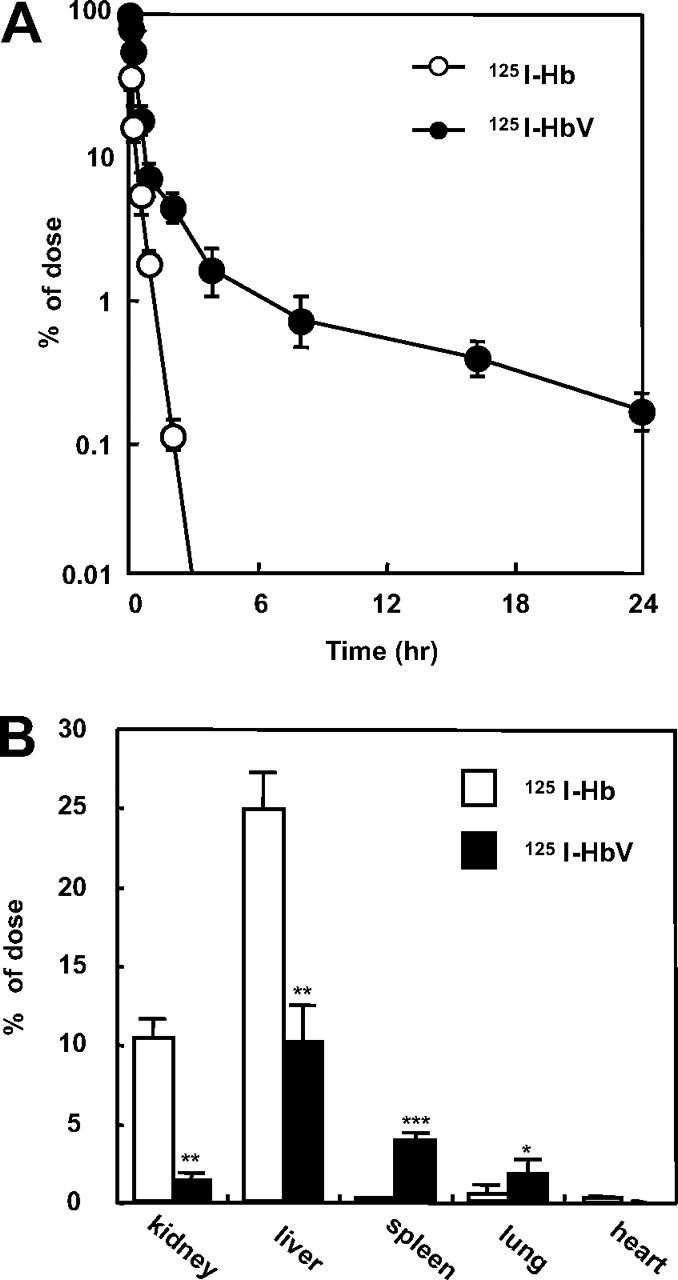
Comparison of Pharmacokinetic Properties between Hb and HbV. The fates of the 125I-Hb and 125I-HbV administered to mice were evaluated as residual TCA-precipitable radioactivity in the plasma. Figure 1A shows the time courses for the plasma concentrations of 125I-Hb and 125I-HbV in mice at a dose of 1 mg of Hb/kg. The blood circulation of HbV was increased notably. According to the pharmacokinetic parameters based on a two-compartment model analysis, the clearance of 125I-HbV was significantly decreased (3.4 ± 0.1 ml/h) compared with that of 125I-Hb (12.7 ± 2.1 ml/h, p < 0.001). The half-life (t1/2) and the area under the concentration-time curve (AUC) of 125I-HbV were increased (t1/2, 3.1 ± 1.0 and 0.1 ± 0.1 h, p < 0.01 and AUC, 29.4 ± 9.2 and 7.9 ± 3.9 h · % dose/ml, p < 0.001, for 125I-HbV and 125I-Hb, respectively) with decreasing clearance, whereas the distribution volume (V1) remained unchanged (2.3 ± 0.1 and 2.6 ± 0.3 ml for 125I-HbV and 125I-Hb, respectively). Figure 1B shows the tissue distribution in each organ at 3 min after an injection of 125I-Hb or 125I-HbV. As expected, 125I-Hb was distributed mainly in the kidney and liver. In contrast, the distribution of 125I-HbV was significantly decreased, whereas its distribution in the spleen and lung was increased significantly.

A, time course for the plasma level of 125I-Hb (○) and 125I-HbV (•) after administration to mice. ddY mice received a single injection of 125I-Hb or 125I-HbV from the tail vein at a dose of 1 mg/kg. Blood was collected from the inferior vena cava under ether anesthesia, and a plasma sample was obtained. Each point represents the mean ± S.D. (n = 3–6). B, tissue distributions of 125I-Hb (□) and 125I-HbV (▪) at 3 min after administration to mice. ddY mice received a single injection of 125I-Hb or 125I-HbV from the tail vein at a dose of 1 mg/kg. At 3 min after injection, each organ was collected. Each bar represents the mean ± S.D. (n = 3–6). *, p < 0.05, **, p < 0.01, and ***, p < 0.001 versus 125I-Hb.
Dose Dependence of HbV Pharmacokinetics.Table 1 shows the pharmacokinetic parameters for 125I-HbV administered to mice at doses of 1, 10, 200, and 1400 mg of Hb/kg. As the dose was increased, t1/2β was increased and clearance was decreased. The AUC was increased in proportion to t1/2β. Table 2 shows the dose-dependent CLuptake in each organ. CLuptake in liver and spleen was much higher than that in the other organs. As the dose was increased, CLuptake in liver and spleen was decreased. These results suggested that the distribution of HbV to the liver and spleen, which contained the majority of HbV, was saturated at higher doses.
Dose-dependent pharmacokinetic parameters of HbV after administration of 125I-HbV in mice
All mice received a single injection of 125I-HbV (1, 10, 200, and 1400 mg of Hb/kg) containing 5% rHSA. At each time (0.05, 0.5, 1, 2, 4, 6, 8, 12, and 24 h) after the 125I-HbV injection, blood samples were collected from the inferior vena cava, and a plasma sample was obtained. Each parameter was calculated by MULTI using a two-compartment model. Values are mean ± S .D. (n = 3–6).
Dose-dependent uptake clearance of HbV in liver, spleen, kidney, lung, and heart after 125I-HbV administration in mice
All mice received a single injection of 125I-HbV (1, 10, 200, and 1400 mg of Hb/kg) containing 5% rHSA. The uptake clearance for each organ was calculated by integration plot analysis at designated times from 1 to 30 min after injection. Values are mean ± S.D. (n = 3–6).
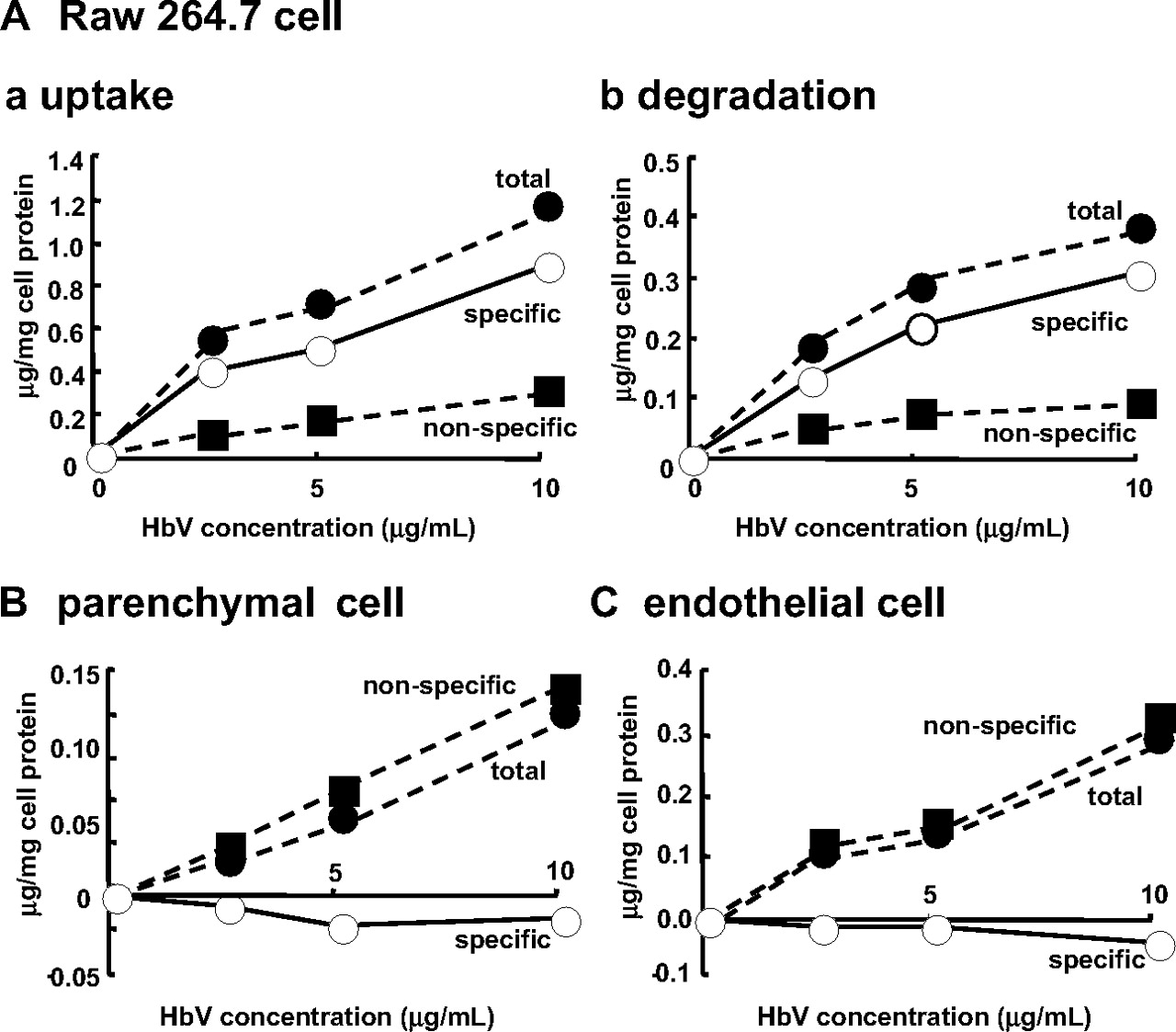
Cell Assay. It is well known that liposomes are scavenged and degraded by the mononuclear phagocyte system (MPS), such as Kupffer cells or macrophages (Kiwada et al., 1998). Because HbV were distributed mainly in the liver and spleen, where the MPS is localized, we examined the issue of whether HbV are scavenged and degraded by MPS using RAW 264.7 cells, which have been used as an alternative to Kupffer cells. In these experiments, primary parenchymal and endothelial cells from mouse livers were used as a control. As shown in Fig. 2, the specific uptake and degradation of 125I-HbV were observed only in the RAW 264.7 cells and not in primary parenchymal and endothelial cells. This in vitro study supports the fact that HbV are captured by Kupffer cells in the liver and by macrophages in the red pulp zone of the spleen, as reported previously in an in vivo study (Sakai et al., 2001).
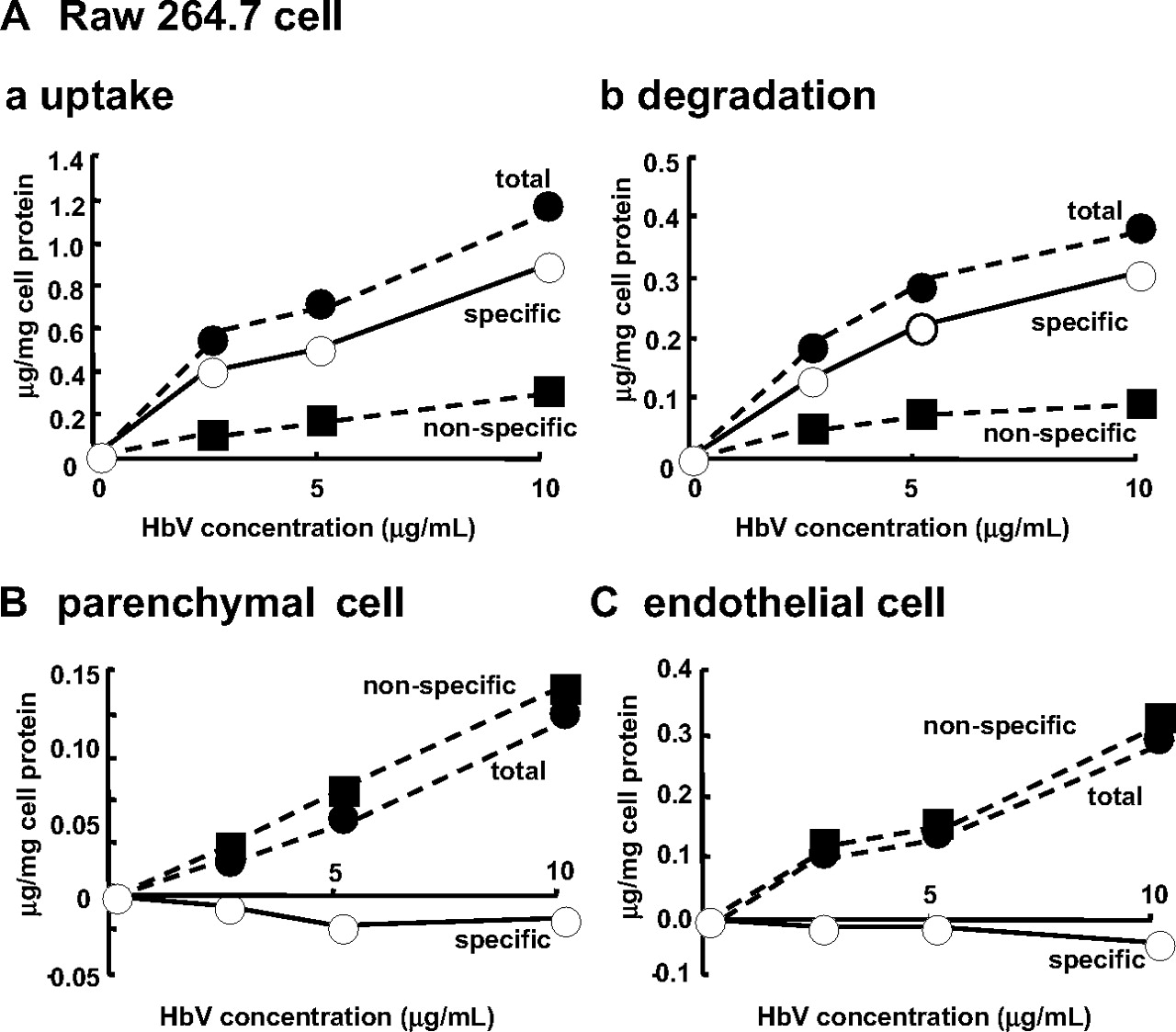
A, dose-dependent endocytic uptake (a) and degradation (b) of 125I-HbV by RAW 264.7 cells. B and C, endocytic uptake of 125I-HbV by parenchymal cells and endothelial cells, respectively. RAW 264.7, primary parenchymal, and endothelial cells were incubated at 37°C for 6 h with the indicated concentration of 125I-HbV in the presence (▪) or absence (•) of 50-fold unlabeled HbV. Specific uptake or degradation (○) was calculated by subtracting the nonspecific values from the total values. Results are the mean ± S.D. of three separate experiments.
Pharmacokinetics of HbV Components in Mice. To investigate the pharmacokinetics of each HbV component, Hb, enclosed in HbV, was radiolabeled with 125I (125I-HbV) or cholesterol, in the lipid component vesicles of HbV, was radiolabeled with 3H (3H-HbV). As shown in Fig. 3 and Table 3, similar plasma concentration curves and pharmacokinetic parameters of 125I-HbV were observed for 3H-HbV. Furthermore, after administration of nonlabeled HbV to mice, the time course for the plasma iron concentration curve derived from HbV was consistent with the plasma concentration curves for both labeled HbV (Fig. 3). These data indicate that HbV is likely to maintain an intact structure in the blood circulation for periods of up to 24 h after injection. We also examined the issue of whether the injected HbV had any influence on the production of reactive oxidative species because, if free iron were released from vesicles, it would enhance the production of ROS by the Fenton reaction, as has been previously demonstrated (Anraku et al., 2004, 2008). To quantitatively evaluate the extent of oxidative stress in the blood circulation, we monitored the ratio of the mercapt-form (nonoxidized form) to the nonmercaptform (oxidized form) of serum albumin, which serves as a marker of oxidative stress in the circulatory system (Kadowaki et al., 2007; Shimoishi et al., 2007). No significant differences in these ratios were found between HbV and the saline administration groups for periods of up to 7 days after administration (data not shown).
Pharmacokinetic parameters of HbV after administration of 125I- and 3H-HbV in mice and rats
All mice and rats received a single injection of 125I-HbV or 3H-HbV at a dose of 1400 mg of Hb/kg containing 5% rHSA. At each time after the injection of 125I-HbV or 3H-HbV, a blood sample was collected from the inferior vena cava (mice) or from the tail vein (rats), and plasma was obtained. Each parameter was calculated by MULTI using a two-compartment model. Values are mean ± S .D. (n = 3–6).

Time course for the plasma level of 125I-HbV (○), 3H-HbV (•), and iron (□) derived from HbV after administration to mice. ddY mice received a single injection of 125I-HbV or 3H-HbV from the tail vein at a dose of 1400 mg of Hb/kg. Blood was collected from the inferior vena cava under ether anesthesia, and a plasma sample was obtained. The iron concentration was calculated from atomic emission spectrometric analysis using an absorbance of 415 nm. Each point represents the mean ± S.D. (n = 3–6).
Moreover, the tissue distribution of 125I-HbV was evaluated using the tissue-to-plasma partition coefficient (Kp). Figure 4 shows the Kp values in organs (kidney, liver, spleen, lung, and heart) 8 h after the administration of 125I-HbV or 3H-HbV to mice. Among these organs, the Kp values for both the liver and spleen reached >1 for both radiolabeled HbV. Therefore, we examined the time course for the tissue distribution in the liver (Fig. 5A) and spleen (Fig. 5B) after the administration of both labeled HbV. At an early time period (Fig. 5, A and B, insets), the time course distributions for 125I-HbV and 3H-HbV were consistent with each other, and the CLuptake values in liver and spleen were also similar (liver, 256 ± 37 and 301 ± 41 μl/h and spleen, 51 ± 6 and 43 ± 12 μl/h for 125I-HbV and 3H-HbV, respectively). However, the radioactive 125I was more rapidly eliminated from each organ, and the activity essentially disappeared within 7 days. On the other hand, the elimination of radioactive 3H was delayed compared with that of 125I and nearly disappeared after 14 days in both liver and spleen. These data indicate that HbV was distributed mainly to liver and spleen in the form of intact HbV and that it was degraded by MPS, followed by different routes of excretion for the internal Hb and the lipid component.
To identify the excretion pathway of HbV, the levels of radioactivity 125I and 3H in urine and feces were measured (Fig. 6). The radioactive 125I was excreted mainly in the urine (84.2 ± 4.1% of ID at 7 days after injection), but was low in feces (5.1 ± 2.3% of ID at 7 days after injection). In addition, neither Hb nor protein urea nor hemoglobinuria was detected in the urine (data not shown). On the other hand, the majority of the radioactive 3H was excreted in the feces (71.1 ± 3.6% of ID at 7 days after injection), and a small portion was excreted into the urine (19.8 ± 3.4% of ID at 7 days after injection). At 7 days after injection, a high level of radioactivity from 3H was detected in plasma lipoprotein and bile, but 125I was not (data not shown).
Pharmacokinetics of the HbV Component in Rats. Because altered pharmacokinetics of liposomes have been reported for different animal species, we carried out a pharmacokinetic analysis in rats as well. The rats showed dose dependence for HbV pharmacokinetics similar to that observed for mice, except that the half-life of HbV was doubled (t1/2 of 8.8 ± 0.7, 11.5 ± 0.3, and 30.6 ± 4.0 h at doses of 10, 200, and 1400 mg of Hb/kg, respectively).
Table 3 shows data for the pharmacokinetic analysis of 125I- and 3H-HbV in rats. Although the pharmacokinetic behaviors of both radiolabeled HbV were similar to those for mice, the maximum hepatic distributions of labeled HbV in rats were decreased by nearly half of those in mice (13.5 ± 0.5 and 17.7 ± 3.0% of ID for 125I-HbV and 3H-HbV, respectively). Moreover, the radioactive 125I was excreted mainly in the urine (77.9 ± 6.1 and 4.8 ± 1.6% of ID at 7 days after injection for urine and feces, respectively). On the other hand, the majority of the radioactivity of 3H was excreted into feces (10.9 ± 4.4 and 66.3 ± 14.8% of ID at 7 days after injection for urine and feces, respectively).
Discussion
The HbV has been developed as an artificial oxygen carrier and has considerable promise for use in clinical settings because of its superb functionality, such as its ability to regulate rheological properties and cardiocirculatory dynamics and its oxygen carrier ability (Izumi et al., 1996; Sakai et al., 2008). In previous studies, cell-free Hb and perfluorocarbon, which had also been developed for use as artificial oxygen carriers, were excluded as possible candidates, because their systemic half-lives were too short (∼0.5–1.5 h) or long (1 year or longer), respectively, for them to effectively function as an optimal oxygen carrier (Savitsky et al., 1978; Nosé, 2004). Therefore, it is necessary to characterize the pharmacokinetic properties of the HbV and its components to demonstrate the efficacy and safety of this preparation because a short half-life leads to a diminished pharmacological effect, whereas a long half-life increases the bioaccumulative potential.
In this study, the half-life of HbV was found to be 30 times higher than that of stroma-free Hb at a dose rate of 1 mg Hb/kg due to decreased distribution to the liver and kidney (Fig. 1). This finding could reflect physicochemical differences, such as diameter, the absence or presence of a membrane structure, and PEG modification between HbV and Hb. In physiological conditions, Hb that is released from ruptured RBCs becomes rapidly bound to haptoglobin (Hp), which promotes CD163 recognition in the liver (Kristiansen et al., 2001). When the Hb concentration exceeds the Hp-binding capacity, unbound Hb is filtered through the kidney (Savitsky et al., 1978). Thus, the reduction in HbV distribution in the liver and kidney could be due to the encapsulation of Hb by the liposome because it may not only suppress the binding of internal Hb to Hp but also inhibit renal glomerular filtration.

The tissue-to-plasma partition coefficient (Kp) of 125I-HbV (□) and 3H-HbV (▪) 8 h after administration to mice. Each bar represents the mean ± S.D. (n = 6).
For HbV, approximately 10% of the dose was distributed to the liver. Because HbV possess a liposome structure, they would be expected to be captured by MPS in the liver and spleen (Kiwada et al., 1998). In fact, as the dose is increased, CLuptake in the liver and spleen was decreased (Table 2), and the half-life was increased (Table 1). In addition, specific uptake and degradation were observed only in macrophage cells but not in parenchymal and endothelial cells (Fig. 2). These results strongly suggest that HbV are scavenged by the MPS, such as Kupffer cells or the red pulp zone, and that this ability became saturated at high doses of HbV. These results are in reasonably good agreement with previous in vivo findings that HbV are taken up by Kupffer cells, the red pulp zone, and mesangial cells (Sakai et al., 2001, 2004a). However, because of increased distribution of HbV into spleen and lung compared with that of free Hb, further study will be needed to demonstrate the effect of HbV administration on the immune and respiratory systems.
In clinical situations such as hemorrhagic shock or an intensive surgical procedure, a massive dose of HbV would be administered to patients. Therefore, it is important to confirm the safety of HbV components from the viewpoint of pharmacokinetics, because Hb molecules can trigger a number of side effects such as renal toxicity and hypertension derived by scavenging endothelium-derived NO (synthesized by NO synthase-3) (Yu et al., 2008) and tissue damage induced by the Fenton reaction mediated by heme (iron) (Balla et al., 2005). In addition, the long-term circulation of high amounts of lipids, especially cholesterol, contributes to cardiovascular and kidney disease (Gröne and Gröne, 2008). In this study, we were able to confirm the safety of HbV components, including Hb, iron, and lipids, based on pharmacokinetic analyses, as evidenced by the following procedures.

Time course for radioactivity in liver (A) and spleen (B) after the administration of 125I-HbV (○) or 3H-HbV (•) at a dose of 1400 mg of Hb/kg to mice. ddY mice received a single injection of 125I-HbV or 3H-HbV from the tail vein at a dose of 1400 mg of Hb/kg. Each point represents the mean ± S.D. (n = 3–6).
First, the findings herein clearly showed that for up to 24 h after injection, the plasma concentration curves of 125I-HbV, 3H-HbV, and iron derived from HbV exhibited similar behaviors (Fig. 3). In addition, CLuptake in liver and spleen was also similar for 125I-HbV and 3H-HbV (see Results). These results indicate that the HbV circulates in the bloodstream as stable, intact vesicles until degraded by the MPS.
Second, nearly 80% of the radioactive 125I is excreted into the urine by 7 days after its injection. This radioactivity is probably due to the degradation of Hb enclosed in liposomes because 125I binds covalently to tyrosine residues of protein (Burger et al., 1983). In fact, neither Hb nor protein urea nor hemoglobinuria was detected in the urine. Moreover, a previous study showed that multiple-dose or a single high-dose treatment of HbV failed to induce any detectable pathological injury in the kidney or any change in arteriolar or venular diameters, which are induced by the dimeric form of Hb or any direct interaction of Hb with NO (Sakai et al., 2001, 2004a; Cabrales et al., 2005). These results indicate that 125I-HbV are degraded by the MPS and that the 125I is excreted in the urine in the form of a degradation product of 125I-Hb derived from HbV.
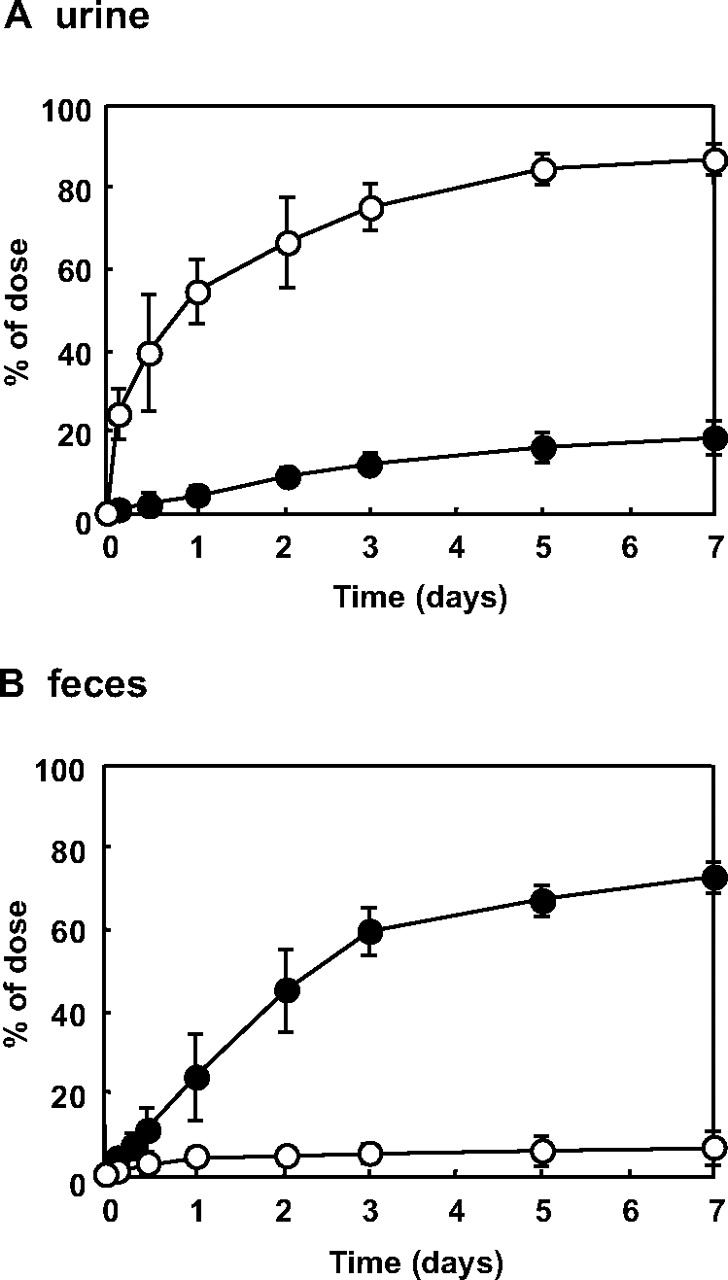
Time course for radioactivity in urine (A) and feces (B) after the administration of 125I-HbV (○) and 3H-HbV (•) to mice. ddY mice received a single injection of 125I-HbV or 3H-HbV from the tail vein at a dose of 1400 mg of Hb/kg. Urine and feces were collected at fixed intervals in a metabolic cage. Each point represents the mean ± S.D. (n = 3–6).
Third, an excess of heme (iron) could cause hemosiderosis and oxidative stress via the Fenton reaction (Buehler and Alayash, 2004). Therefore, the disposition of heme (iron) derived from HbV needs to be clarified. Our study showed that the plasma concentration curve for heme (iron) derived from HbV was similar to that for 125I-HbV and 3H-HbV (Fig. 3). In addition, after an HbV injection, systematic oxidative stress, as estimated by the oxidized albumin ratio, was not increased (data not shown). These results suggest that an excess amount of free heme (iron) derived from HbV was not released in the plasma. In fact, a previous study reported that a daily repeated infusion of HbV (1000 mg of Hb/kg/day) for 14 days had no effect on plasma iron and bilirubin levels. Therefore, phospholipid vesicles for the encapsulation of Hb would be beneficial for heme detoxification through their preferential delivery to the MPS as described in a previous report (Sakai et al., 2004a). Furthermore, deposition of hemosiderin, which is siderosis as heme detoxification, was detected in the liver and spleen after the administration of HbV (2000 mg of Hb/kg), but this deposition completely disappeared after 14 days (Sakai et al., 2001). In the future, it will be necessary to study ferrokinetics to confirm this issue by using Hb with radiolabeled iron (e.g., 55Fe and 59Fe). It will also be interesting to determine whether the deposited iron is effectively used for hematopoiesis after massive blood loss and HbV administration.
Finally, the findings herein show that [3H]cholesterol in HbV was mainly distributed to the liver and spleen, and subsequently excreted into feces approximately 14 days after 3H-HbV injection (Figs. 4 and 5). These results are in good agreement with histopathological examinations, using oil red O staining of liver tissue after an injection of HbV (2000 mg of Hb/kg), which revealed that slight stains were confirmed 3 days after injection and that this staining disappeared within 7 days (Sakai et al., 2001). In addition, Sakai et al. (2004a) found that daily repeated infusion of HbV for 14 days elevated the level of plasma lipid components, one of the risk factors of atherosclerosis, although the effect was temporary. However, cholesterol in the vesicles should reappear in the blood mainly as lipoprotein cholesterol after entrapment in the Kupffer cells and should then be excreted in bile after entrapment of the lipoprotein cholesterol by the hepatocytes (Kuipers et al., 1986). In fact, high levels of radioactivity of 3H in plasma lipoproteins and bile, which are related to carrying and excretion of endogenous cholesterol by physiological pathways, were observed (data not shown). Therefore, it would be desirable to know that cholesterol in the lipid components in HbV behaved the same as endogenous cholesterol after the metabolization of HbV in the MPS.
Conversely, we did not directly examine the disposition of the phospholipid, DPPC, in HbV. Previous reports have shown that phospholipids in the liposome are metabolized in the MPS and reused as cell membranes or are excreted into the bile (Dijkstra et al., 1985; Verkade et al., 1991). Therefore, it is also possible that phospholipids in HbV are metabolized and excreted in the same manner as mentioned above. However, further study will be needed to demonstrate this fact.
Based on the present findings, we propose that the disposition of HbV and its components occurs as follows. After circulating in the form of stable HbV, they are distributed to the liver and spleen, where they are degraded by the MPS. Finally, the enclosed Hb and outer lipid components are eliminated mainly to the urine and feces, respectively, in the same manner as endogenous substances. In addition, our pharmacokinetic study using different animal species enabled us to predict pharmacokinetics in humans. In fact, we reported previously that the half-life of HbV in humans was estimated to be approximately 3 to 4 days by using an allometric equation (Taguchi et al., 2009). The above findings provide further support for the effectiveness and safety of the HbV for use as an oxygen carrier.
Footnotes
-
This work was supported in part by the Ministry of Health, Labour and Welfare, Japan [Health Science Research Grants (Health Science Research Including Drug Innovation)].
-
K.T. and Y.U. contributed equally to this work.
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.109.027094.
-
ABBREVIATIONS: RBC, red blood cell; HbV, hemoglobin vesicle(s); PEG, polyethylene glycol; NO, nitric oxide; 125I-HbV, 125I-labeled hemoglobin vesicle; 3H-HbV, 3H-labeled hemoglobin vesicle; DPPC, 1,2-dipalmitoyl-sn-glycero-3-phosphatidylcholine; rHSA, recombinant human serum albumin; BSA, bovine serum albumin; TCA, trichloroacetic acid; PBS, phosphate-buffered saline; WE, Williams' E medium; FCS, fetal calf serum; AUC, area under the concentration-time curve; MPS, mononuclear phagocyte system; ID, injected dose; Hp, haptoglobin; CLuptake, uptake clearance.
- Accepted April 9, 2009.
- Received February 11, 2009.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}