Article Text

Abstract
Introduction Vitamin D3 supplementation has been reported to prevent lung infections and increase the gene expression of antimicrobial peptides such as cathelicidin. We investigated the effect of vitamin D3 supplementation on the antimicrobial activity of airway surface liquid (ASL) in human subjects. Since smoking can increase the risk of respiratory infections, we also investigated the effect of smoking in the cathelicidin response to vitamin D3 in human airway epithelia in vitro.
Methods This study is a subanalysis of single-centre community-based randomised placebo-controlled double-blind trial. Participants were randomised to receive 1000 international units per day of oral vitamin D3 or identical placebo for 90 days. Blood and ASL samples were collected preintervention and postintervention. 105 participants were originally enrolled, 86 completed the trial, and due to low protein concentration in the samples, 40 participants were finally analysed. Our primary outcome was ASL antimicrobial activity. We also considered secondary outcomes including changes in serum concentration of 25-hydroxyvitamin D3 (25(OH)D3), 1,25-hydroxyvitamin D3, calcium and parathyroid hormone (PTH). In addition, we studied the effect of cigarette smoke extract (CSE) exposure to primary human airway epithelial cell cultures on the gene expression of cathelicidin in response to vitamin D3 and expression of CYP27B1 (1-alpha hydroxylase), responsible for vitamin D3 activation.
Results Vitamin D3 supplementation significantly increased both ASL antimicrobial activity and serum concentration of 25(OH)D3. In a subgroup analysis, we found that smokers did not increase their baseline antimicrobial activity in response to vitamin D3. Exposure to CSE on human airway epithelia decreased baseline CYP27B1 gene expression and cathelicidin response to 25(OH)D3.
Conclusion Vitamin D3 supplementation for 90 days increases ASL antimicrobial activity. Data from this preliminary study suggest that smoking may alter the ability of airway epithelia to activate vitamin D3 and increase the gene expression of cathelicidin antimicrobial peptide.
Trial registration number NCT01967628; Post-results.
- Airway Epithelium
- Innate Immunity
- Bacterial Infection
- Respiratory Infection
- Tobacco and the lung
This is an Open Access article distributed in accordance with the terms of the Creative Commons Attribution (CC BY 4.0) license, which permits others to distribute, remix, adapt and build upon this work, for commercial use, provided the original work is properly cited. See: http://creativecommons.org/licenses/by/4.0/
Statistics from Altmetric.com
Strengths and limitations of this study
We used airway surface liquid samples obtained through bronchoscopy from the same human subjects before and after interventions.
The samples used are from a double-blind, randomised, placebo-controlled trial.
We had a small number of samples in the subgroup analysis (smoking vs non-smoking group) that will require further confirmation in larger studies.
Introduction
Vitamin D3 is a liposoluble vitamin naturally present in very few dietary sources and generated by the skin when exposed to sunlight. Despite supplementing food with vitamin D, a great proportion of the world’s population is still deficient.1–3 Vitamin D deficiency is found more often in smokers with accelerated lung function decline and in patients with severe chronic obstructive pulmonary disease (COPD).4–8 One of the proposed mechanisms for lung function decline and development of chronic respiratory diseases is the occurrence of airway infections.9 10 Since low vitamin D3 levels increase susceptibility to respiratory infections in both smokers and non-smokers,11–14 it is critical to investigate the effect of vitamin D supplementation on airway antimicrobial activity in both populations.
Vitamin D3 in the bloodstream is hydroxylated by the liver into 25-hydroxyvitamin D3 (25(OH)D3). This form is used conventionally to measure vitamin D3 sufficiency. Thereafter, the 1α-hydroxylase, constitutively active in the kidney, activated macrophages and in airway epithelia converts 25(OH)D3 into 1,25-dihydroxyvitamin D3 (1,25(OH)2D3).15 The active form of vitamin D3 binds to the intracellular vitamin D receptor to activate vitamin D response elements to modulate the gene expression of multiple pathways involved in serum calcium metabolism and bone remodelling.16 It also upregulates the expression of antimicrobial peptides (AMPs) present in the airway surface liquid (ASL), such as human cathelicidin (LL-37) and human beta defensins (hBDs) secreted by airway epithelia and immune cells.15 17–19 AMPs can kill pathogens within minutes as a first line of defence and their impairment is associated with the development of airway disease.20 Various mechanisms can impair AMP activity, such as: decreased production, decreased pH, increased ionic strength, increased host and pathogen proteases and increased polymerisation of AMPs with DNA/filamentous-actin.21–26
We hypothesised that vitamin D3 supplementation would increase human ASL antimicrobial activity in vivo. To test this hypothesis, we investigated the antimicrobial activity of biobanked ASL samples collected through bronchoscopy of a cohort of smoking and non-smoking participants that randomly received either 1000 international units (IU) of vitamin D3/day or matching placebo for 90 days.27 We also investigated the effect of smoking in the airway response to vitamin D3 in vivo and in vitro.
Methods
Study design
This study design has been previously published.27 Briefly, this is a single-centre, community-based, randomised, placebo-controlled double-blind trial. This pilot study investigated the effects of vitamin D3 supplementation on the innate immunity of the lung. The University of Iowa Hospital and Clinics Institutional Review Board approved the trial (IRB# 200607708). Written consent was obtained from all the subjects. This trial is registered on ClinicalTrials.gov (NCT01967628).
Participants
We recruited subjects 18–60 years old from the Iowa City community. We excluded participants if they were taking vitamin supplements within the previous 3 months, had any history of positive tuberculin test, pneumonia within 3 years, airway infection or antibiotic use within 6 weeks or vaccination within 1 month of enrolment. Additional exclusion criteria included any prescription medication except: oral contraceptives, topical medications, selected antidepressants, levothyroxine, acid reflux treatment, over-the-counter antihistamine or sleep aids. Participants were also excluded if they had one of these conditions: pregnancy, breastfeeding, asthma, diabetes, heart disease or allergy to lidocaine. We also excluded subjects that did not complete their second visit or if ASL collection was not possible due to technical limitations during bronchoscopy (figure 1).
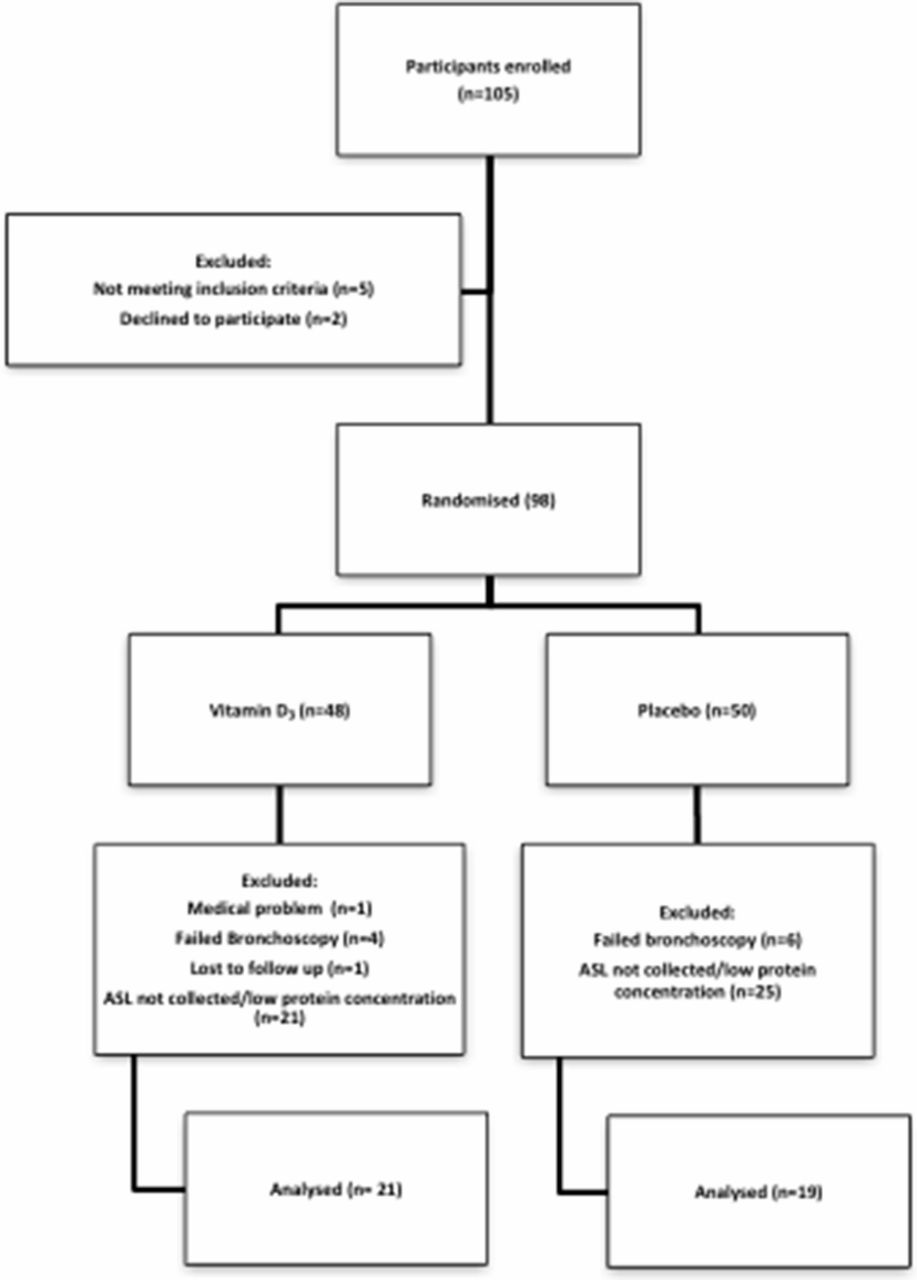
Participant flow diagram.
Interventions and randomisation
After informed consent was signed and pregnancy was excluded, the subject’s blood was drawn to assess 25(OH)D3, 1,25(OH)2D3, parathyroid hormone (PTH) and calcium serum concentration. Thereafter, a bronchoscopy was performed at the University of Iowa Hospitals and Clinics (Iowa City, Iowa, USA) as previously described.27 To collect the ASL, three sponges were placed, one at a time, on the right main stem bronchus or bronchus intermedius for 30 s and then eluted with 1 mL of isotonic saline into a 1.5 mL tube. The tube was vortexed for 1 min and the sponges removed. The samples were stored at −80°C. After collections, participants were randomised using a computer-generated list to receive either vitamin D3 (1000 IU/day, 90 000 IU total) or matching placebo. Study participants, the clinical research team and the sponsoring agency were blinded to intervention assignments.
Outcomes
The primary outcome analysed for this subanalysis was ASL antimicrobial activity preintervention and postintervention (vitamin D3 or placebo). We considered secondary outcome changes in serum concentration of 25(OH)vitamin D3 (25(OH)D3), 1,25(OH)2D3, calcium and PTH. In a post hoc analysis, we also investigated the difference between the smoking and non-smoking groups in response to vitamin D3.
ASL protein concentration
Briefly, aliquots were thawed at 4°C and total protein concentration was measured by a Bradford assay, interpolating samples to a bovine serum albumin standard curve. We adjusted sample concentrations with normal saline to 100 µg/mL of total protein. This threshold was selected prior the antimicrobial activity assay based on preliminary antimicrobial activity studies to minimise sample dilution and yet still be able to include the majority of the subjects.
ASL antimicrobial activity
We added 10 µL of protein-corrected ASL (100 µg/mL) or protein uncorrected into a 96-well plate. Then, samples were challenged with 10 µL of a solution containing bioluminescent Staphylococcus aureus Xen 29 (~5×106 colony forming units) injected by the precision peristaltic pump in the luminometer. After 4 min, we measured relative light units (RLU) as a surrogate for live bacteria as previously described.21 To minimise observer bias, we randomised the samples onto the plate. The luminometer injected the bacteria and measured RLU automatically, and the conditions were not revealed until the experiment was completed.
Macrophage microarray
RNA preparation, quality analysis and microarray analysis were done as previously described.28 Measurements of genome-wide macrophage messenger RNA expression and analysis were done as previously described27 (Gene Expression Omnibus (GEO) accession # GSE56583).
Human airway epithelia exposure to cigarette smoke extract
Differentiated primary human airway epithelia were cultured at the air liquid interface as previously described.29 We switched cultures to minimum essential media, with and without 25(OH)D3 (10-7 M) or 1,25(OH)2D3 (10−7 M) in the presence or absence of 3% cigarette smoke extract (CSE).7 15 We changed the media every day for the duration of the experiment and analysed by quantitative (q)PCR the relative gene expression of CYP27B1 which codes for 1α-hydroxylase and cathelicidin antimicrobial peptide (CAMP) which codes for the cationic antimicrobial peptide LL-37. First, we exposed cells to 25(OH)D3 or 1,25(OH)2D3 or CSE alone for 4 days and quantified the relative gene expression of CAMP and CYP27B1 (n=4). In separate experiments, we analysed the response to 25(OH)D3 (n=8) or 1,25(OH)2D3 (n=5) in epithelia exposed overnight to CSE or control and analysed the relative gene expression of CAMP by qPCR.
Compliance
In the vitamin D3 group, 46% of subjects were compliant and 39% had residual doses. The mean number of missed capsules was approximately 6.5 capsules (range 1–15) for an average of total missed vitamin D3 of 6500 IU per subject. Four treated subjects did not bring back their pill bottles for compliance analysis. In the placebo group, 43% of subjects completed full therapy and 53% had missed doses. The mean number of missed capsules was 8.8 (range 1–30). One placebo subject did not bring back the pill bottle for analysis.
Statistical analysis
Data are expressed as mean ± SEM For ASL antimicrobial activity, we used raw RLU and for the LL-37 experiments we normalised data as per cent of control using this formula:

To determine the statistical significance between preintervention and postintervention (vitamin D3 and placebo), we used paired t-tests. We used multiple comparison analysis of variance (ANOVA) and Kruskal-Wallis to compare three or more conditions in the antibody treated ASL and LL-37. We also used Pearson test to calculate correlation coefficients. We did not exclude improbable data. Data analysis was performed using GraphPad Prism V.6.00 (GraphPad Software, California, USA). Power analysis were performed using G*Power V.3.1.9.2 (Germany).
Results
Participant demographics
We first analysed the differences in demographics between subjects receiving vitamin D3 or placebo (table 1). The vitamin D3 group had slightly more percentage of white, male and non-smoking participants compared with the placebo group. We found no significant differences in the age and number of pack-years in smokers between vitamin D3 and placebo groups. We also analysed the differences in serum concentration of 25(OH)D3, 1,25(OH)2D3, calcium and PTH before intervention between the vitamin D3 and placebo groups and found no significant differences (table 2).
Comparison of patient characteristics at baseline by treatment group
Treatment effect on several serum markers of vitamin D metabolism
Effect of vitamin D3 on serum levels of canonical vitamin D3 metabolites
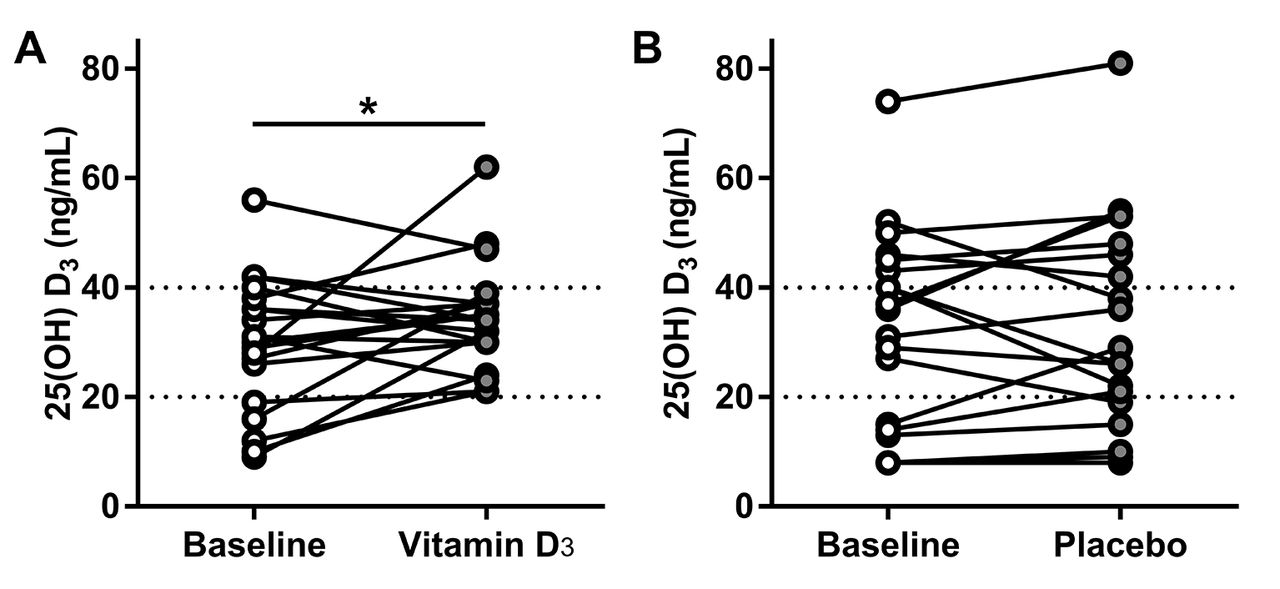
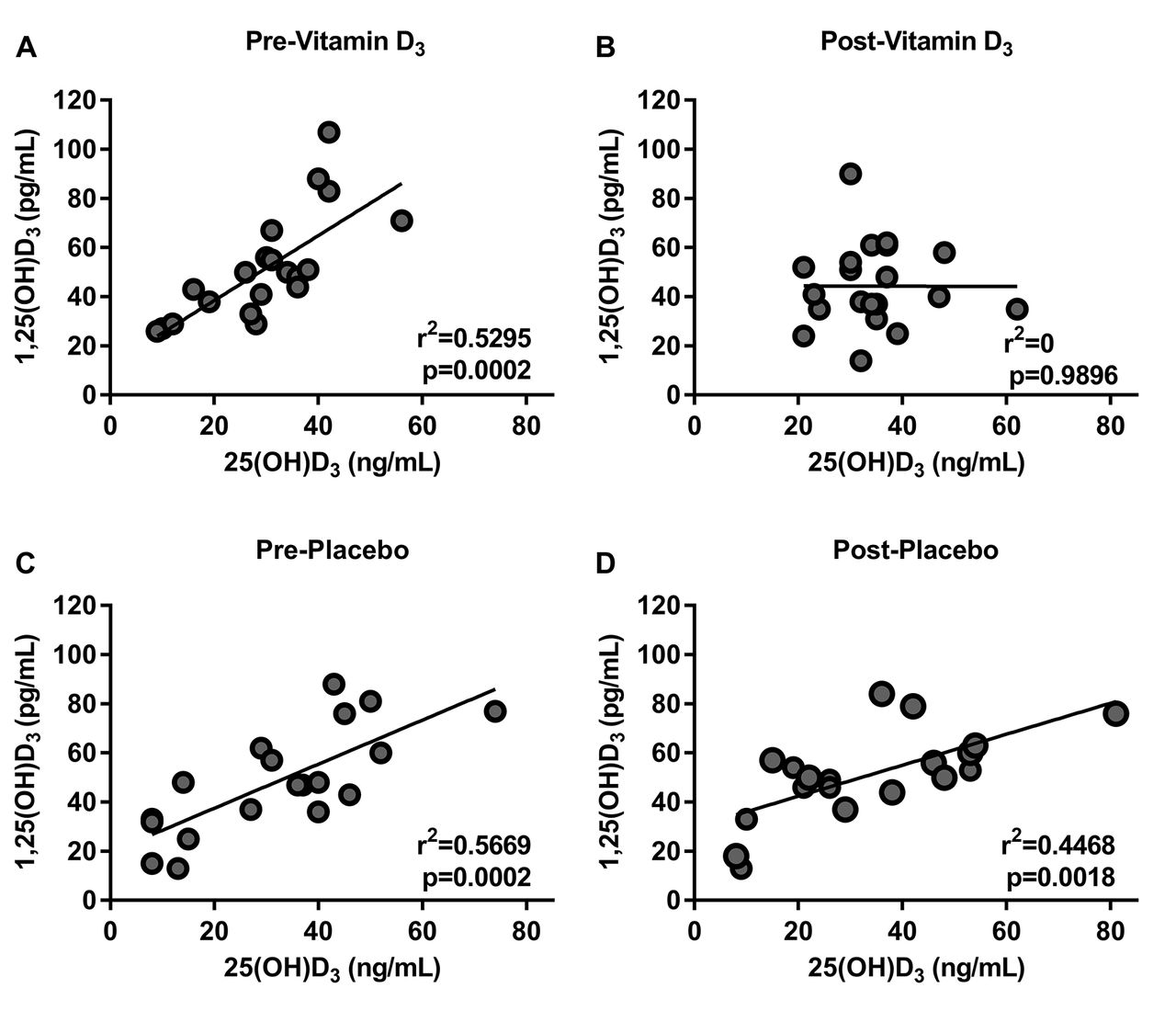
We investigated the effect of vitamin D3 or placebo on the serum concentration of several canonical vitamin D metabolites such as: 25(OH)D3, 1,25(OH)2D3, calcium and PTH (table 2). We found that most of the participants had 25(OH)D3 levels less than 40 ng/mL and that only participants taking vitamin D proportionally increased their baseline 25(OH)D3 levels (figure 2). There was no statistically significant difference in the serum concentration of the other metabolites measured in either intervention (table 2). We also found that there was a positive correlation between 25(OH)D3 and 1,25(OH)2D3 at baseline in both groups. After the intervention, this correlation persisted in the placebo group but it was absent after vitamin D3 supplementation consistent with a biological effect of vitamin D3 (figure 3).

Treatment effect on serum concentration of 25(OH)D3. (A) Effect of vitamin D3 supplementation on serum concentration of 25(OH)D3 (*p=0.0461 by ratio paired t-test). (B) Effect of placebo supplementation on the serum concentration of 25(OH)D3. Dotted line at 20 ng/mL represents deficiency and above 40 represents optimal levels.
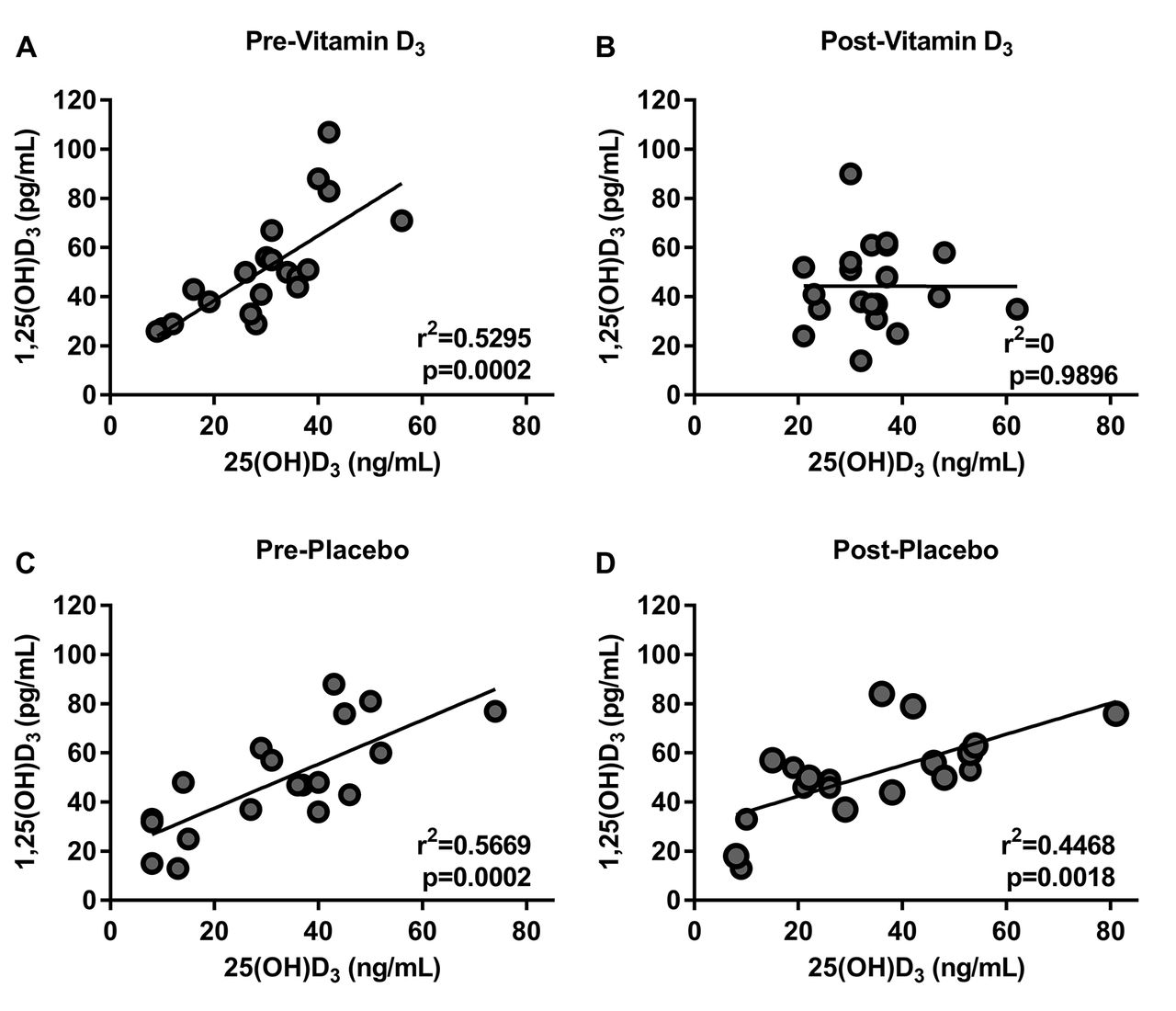
Vitamin D3 supplementation decreased the correlation of 25(OH)D3 to 1,25(OH)2D3. Pearson correlation of 25(OH)D3 and 1,25(OH)2D3 at baseline (A) previtamin D3, (B) postvitamin D3, (C) preplacebo, (D) and postplacebo. At baseline, there was a good correlation between the levels of 25(OH)D3 and 1,25(OH)2D3 in all groups (A and B). After vitamin D3 supplementation, the correlation disappears, while the placebo maintained this positive correlation (C and D).
Vitamin D3 increases ASL antimicrobial activity
We investigated the effect of vitamin D3 supplementation on ASL antimicrobial activity using a bioluminescent bacterial challenge. We challenged ASL samples with bioluminescent bacteria and measured live bacteria by RLU after 2 min as a surrogate of antimicrobial activity. We interpreted a reduction in live bacteria after challenge in RLU as increased antimicrobial activity. We found that subjects taking vitamin D3 supplementation for 90 days had significantly lower live bacteria in RLU after challenge compared with their baseline (95% CI of the difference −1122 to −40.37; p=0.0365 by paired t-test) (figure 4A). Placebo treatment did not significantly decrease the number of live bacteria after challenge compared with baseline (95% CI of the difference −1170 to 198.5; p=0.1532 by paired t-test) (figure 4B). These results suggest that vitamin D3 supplementation for 90 days increases baseline ASL antimicrobial activity.

Vitamin D3 increased ASL antimicrobial activity. ASL antimicrobial activity assay. (A) ASL antimicrobial activity at baseline and after vitamin D3 (Baseline vs vitamin D mean±SD, 5980±1201 vs 5399±855.8; 95% CI of the difference −1122 to −40.37; *p=0.0365 by paired t-test). (B) ASL antimicrobial activity at baseline and after placebo. Placebo treatment did not increase the ASL antimicrobial activity compared to baseline (baseline vs placebo Mean±SD, 6155±1130 vs 5670±1089; 95% CI of the difference −1170 to 198.5; p=0.1532 by paired t-test). ASL, airway surface liquid; LL-37, human cathelicidin.
25(OH)D3 deficiency was associated with lower ASL antimicrobial activity
We hypothesised that subjects with serum vitamin D3 deficiency (<20 ng/mL) would have lower ASL antimicrobial activity compared with non-deficient subjects (≥20 ng/mL). To investigate the effect of vitamin D3 deficiency on ASL antimicrobial activity, we paired RLU values in the samples with their respective serum 25(OH)D3 regardless of allocation group (vitamin D3 or placebo) or whether they were baseline or postintervention samples. Thereafter, we compared the amount of live bacteria after challenge in the ASL samples with a clinically deficient 25(OH)D3 serum concentration to the samples with non-deficient levels. We found that ASL samples associated with vitamin D3 deficiency had significantly more live bacteria after challenge compared with non-deficient subjects (6394±289, n=16 vs 5646±129, n=64 respectively, p=0.0136 by unpaired t-test). These data suggest that vitamin D3 deficiency is associated with a decrease in ASL antimicrobial activity.
LL-37 antibody inhibits ASL antimicrobial activity
We hypothesised that ASL antimicrobial activity is dependent on LL-37 activity. Since we were limited by volume and concentration of the samples, we took an indirect approach to investigate this hypothesis. We used an LL-37 antibody to neutralise the effect of cathelicidin, one of the antimicrobial peptides regulated by the vitamin D response elements, as previously described.23 30 31
When we coincubated increasing concentrations of LL-37 antibody with synthetic LL-37, we observed inhibition of the antimicrobial activity in a concentration-dependent manner compared with control antibody (goat IgG, antimouse IgG) (figure 5A). We also tested the effect of increasing doses of LL-37 antibody on seven random preintervention ASL samples from non-smokers, resulting in decreased ASL antimicrobial activity in a concentration-dependent manner (figure 5B). When we treated ASL samples with control antibody at maximal concentration, we observed no effect compared with ASL without the antibody at 4 min after bacterial challenge (figure 5C). These results suggest that LL-37 antibody can inhibit ASL antimicrobial activity.

LL-37 antibody inhibits both LL-37 and ASL antimicrobial activity. (A) Antimicrobial activity assay of synthetic human LL-37 coincubated with increasing concentration of LL-37 antibody or IgG control (n=3). (B) Antimicrobial activity of ASL samples coincubated with increasing concentrations of LL-37 antibody (n=6). (C) ASL antimicrobial activity of ASL samples coincubated with LL-37 antibody (70 nM) and IgG (70 nM) as control (N=6; *p=<0.05 by multiple comparisons ANOVA). (D) Antimicrobial activity of ASL samples before and after vitamin D3 supplementation treated with an LL-37 antibody (N=21; *p=0.0196 by paired t-test). ANOVA, analysis of variance; ASL, airway surface liquid; LL-37, human cathelicidin.
Anti-LL-37 blocks the vitamin D3-induced improvement of ASL antimicrobial activity
We hypothesised that LL-37 antibody would block the increased antimicrobial activity of the ASL samples from non-smokers postvitamin D3 supplementation. When we coincubated ASL samples previtamin D3 and postvitamin D3 with LL-37 antibody (70 nM), the increased antimicrobial activity was significantly reversed (figure 5D).
Vitamin D3 does not increase cathelicidin gene expression in alveolar macrophages
AMPs in ASL can come from two main sources: inflammatory cells such as macrophages and airway epithelial cells. Since it has been reported that vitamin D3 increases LL-37 in both human macrophages and airway epithelia,15 32 we investigated the change in CAMP gene expression in human alveolar macrophages from smokers and non-smokers treated with vitamin D3. We found no significant difference in either smokers or non-smokers (fold change −1.05, p=0.33 in non-smokers and fold change −1.15, p=0.16 in smokers). These results suggest that airway epithelia might be responsible for the increase in ASL antimicrobial activity due to vitamin D3.
25(OH)D3 and 1,25(OH)2D3 increase airway CAMP gene expression
Since we did not find an increase in CAMP gene expression in macrophages, we investigated the effect of vitamin D3 exposure in primary human airway epithelial cells cultured at the air–liquid interface. As expected, supplementing the media with 25(OH)D3 and 1,25(OH)2D3 increased the relative gene expression of CAMP. In addition, vitamin D3 supplementation decreased CYP27B1, which encodes 1-alpha hydroxylase, the enzyme that converts vitamin D3 into its active form (figure 7A,B).15 This decrease in CYP27B1 is consistent with a self-regulatory loop that decreases excessive vitamin D3 activation.15 33

Human airway epithelia exposed to CSE had decreased gene expression of CAMP in response to 25(OH)D3 and decreased CYP27B1. (A and B) Gene expression of CYP27B1 and CAMP by qPCR of primary human airway epithelia exposed to 25(OH)-D3 (A) and 1,25(OH)2D3 (B). Both forms of vitamin D3 increased CAMP relative gene expression and decreased CYP27B1. (C) Gene expression of CAMP in response to 25(OH)D3. Exposure to CSE blunted the increase in CAMP in response to 25(OH)D3 (*p=0.0398 by paired t-test). (D) Gene expression of CAMP in response to 1,25(OH)2D3. Exposure to CSE did not affect the increase in CAMP in response to 1,25(OH)2D3. (E) Gene expression of CYP27B1 qPCR of primary human airway epithelia exposed to CSE. Exposure to CSE decreased the relative gene expression of CYP27B1. CAMP, cathelicidin antimicrobial peptide; CSE, cigarette smoke extract; qPCR, quantitative PCR.
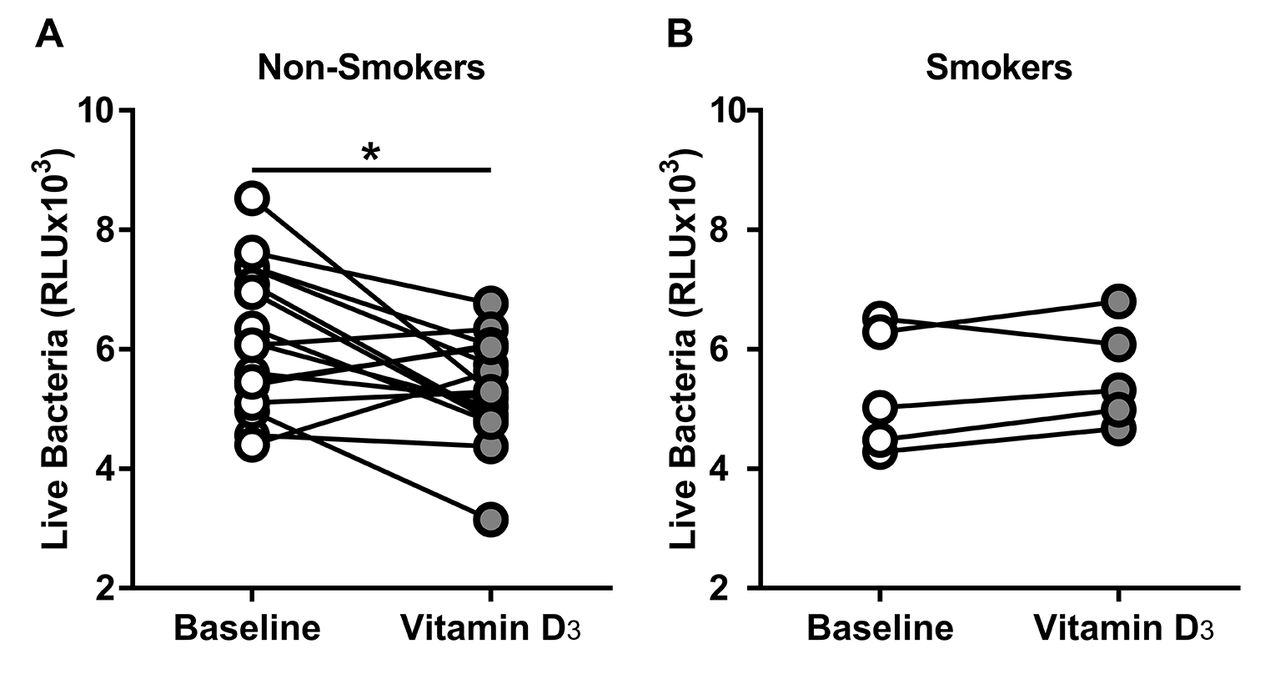
Smokers did not increase ASL antimicrobial activity
In a post hoc analysis, we investigated the effect of smoking in the antimicrobial activity of the ASL. When we analysed the ASL from smokers and the non-smokers, only the non-smokers significantly decreased the number of live bacteria compared with their baseline (baseline vs vitamin D3 in non-smokers) (figure 6A). Although the smokers had a small sample size, we did not observe any difference (figure 6B). Using the data from the non-smokers that received vitamin D3 we determined an effect size of 0.8462, and using a power of 0.8 and alpha of 0.05, we calculated an a priori sample size of at least 14 subjects to detect a difference in the smokers.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Vitamin D3 increased ASL antimicrobial activity in non-smokers. ASL antimicrobial activity assay. (A) Non-smokers ASL antimicrobial activity at baseline and after vitamin D3. Non-smokers significantly increased their baseline antimicrobial activity (baseline vs vitamin D3 mean±SD, 6186±1205 vs 5344±874.3; 95% CI of the difference −1502 to −181.9; *p=0.0159 by paired t-test). (B) Smokers ASL antimicrobial activity at baseline and after vitamin D3. Smokers did not increase their ASL antimicrobial activity (baseline vs vitamin D3 mean±SD, 5321±1029 vs 5575±863; 95% CI of the difference −235.1 to 743.2; p=0.2227 by paired t-test). ASL, airway surface liquid.
CSE decreased CAMP gene expression response to vitamin D3
Since smokers did not improve their baseline antimicrobial activity when supplemented with vitamin D3, we investigated the effect of CSE exposure on CAMP gene expression in response to 25(OH)D3 in vitro. We found that airway epithelial cells exposed to CSE and supplemented with 25(OH)D3 had a smaller increase in CAMP gene expression compared with unexposed cells (figure 7C). We then investigated if supplementing 1,25(OH)2D3 would overcome this decreased response in CAMP gene expression in CSE-exposed human airway epithelial cells. We found a similar increase in CAMP gene expression in both CSE-exposed and unexposed human airway epithelial cells in response to 1,25(OH)2D3 (figure 7D). We conclude that CSE exposure decreases cathelicidin gene expression in response to 25(OH)D3. This can be overcome by supplementing 1,25(OH)2D3, suggesting that CSE might affect the activation of vitamin D3.
CSE decreases airway CYP27B1 gene expression
We hypothesised that exposure to cigarette smoke decreases the gene expression of 1-alpha hydroxylase, responsible for the activation of 25(OH)D3. To test this hypothesis, we exposed human airway epithelial cells to CSE and analysed the gene expression of CYP27B1 by qPCR. We found that exposure to CSE decreased CYP27B1 gene expression compared with unexposed cells (figure 7E). This result suggests that cigarette smoke exposure might decrease the activation of vitamin D3.
Discussion
Prevention of respiratory infections by vitamin D3 supplementation is controversial. Some reports show that vitamin D3 supplementation reduces the risk of developing respiratory infections34 while others do not show such protection.35 To our knowledge, this is the first in vivo study demonstrating that vitamin D3 supplementation increases baseline human ASL antimicrobial activity using a standardised sample and inoculum.
In our cohort, vitamin D3 supplementation for 90 days increased 25(OH)D3 serum concentration and increased ASL antimicrobial activity. To explain the later result, we presume that as calcium concentration increases several negative feedback loops tightly regulate vitamin D3 activation. For example, increased 1,25(OH)2D3 concentrations decreases 1-α hydroxylase and increases 24-hydroxylase, which is responsible for inactivation of both 25(OH)D3 and 1,25(OH)2D3.33
In addition, we found that ASL samples matched with a deficient vitamin D3 serum concentration had significantly lower antimicrobial activity compared with samples paired with sufficient vitamin D3. These results are consistent with epidemiological studies that report that low vitamin D3 levels increase the susceptibility to respiratory infections.11–14
We speculate that vitamin D3 increases the ASL antimicrobial activity by upregulating gene expression of antimicrobial peptides like cathelicidins.15 17–19 Since we did not find any difference in CAMP gene expression in macrophages, we hypothesised that the airway epithelium was responsible for the increase in ASL antimicrobial activity due to vitamin D3. Since we were limited by the volume and concentration of the samples, we took an indirect approach to confirm our hypothesis. As a proof of concept, we used an LL-37 antibody to neutralise the effect of LL-37 in the ASL.23 30 We found that the improvement in ASL antimicrobial activity after vitamin D3 supplementation was reversed when the ASL was treated with an LL-37 antibody. This effect can be in part due to both functional inhibition of this antimicrobial peptide and/or decreased synergism between LL-37 and other antimicrobial peptides such as hBDs.22 Future studies should consider sampling more volume of ASL to be able to measure directly AMPs. Nonetheless, He et al found that Vitamin D3 supplementation for 14 weeks increased the concentration of cathelicidin in both plasma and saliva in healthy subjects.36
We also did a subgroup analysis on the subjects who were smokers, and found compelling results that suggest that they did not increase their ASL antimicrobial activity after vitamin D3 supplementation. We acknowledge that our sample size was small and we might be underpowered to detect the difference. However, our in vitro study showing that human airway epithelial cells exposed to CSE in vitro had decreased cathelicidin gene expression in response to 25(OH)D3 strongly supports the feasibility of our in vivo finding, despite lack of power. In addition, it has been reported that cigarette smoke exposure is negatively correlated with LL-37 in human saliva.37 Furthermore, patients with COPD who have high risk of frequent exacerbations also have decreased plasma levels of LL-37 compared with control subjects.38 These results warrant further evaluation in larger sample sizes.
Our in vitro results also suggest that one of the mechanisms responsible for this finding is that cigarette smoke reduces gene expression coding for the enzyme responsible for converting vitamin D3 into its active form. This is consistent with a prior report that exposure to cigarette smoke was associated with impaired ability of human airway epithelia to hydroxylate 25(OH)D3 to 1,25(OH)2D3.7 When we used the active form of vitamin D3, we found a similar increase in cathelicidin gene expression regardless of CSE exposure. Heulens et al also reported that human macrophages exposed to CSE had a similar increase in cathelicidin gene expression in response to 1,25(OH)2D3compared with unexposed cells.39
Given that cigarette smoke exposure (1) decreased cathelicidin gene expression in response to 25(OH)D3, (2) decreased CYP27B1 gene expression and (3) did not affect the cathelicidin gene expression response to active vitamin D3, we propose that one mechanism by which CSE impairs airway innate immunity is by decreasing local vitamin D3 activation.
Another plausible explanation for decreased response to vitamin D3 in the smokers is different processing of hCAP18, the parent molecule of LL-37 before being processed or LL-37 itself. Smoking increases the concentration of proteolytic proteins such as cathepsin D and neutrophils elastase.40 41 Proteolysis by these enzymes can decrease LL-37 in the airways and its antimicrobial activity.23
Furthermore, although cathelicidin is a relevant antimicrobial peptide, other antimicrobials that have a vitamin D response element like hBDs might be also increased by vitamin D3 supplementation.42 Herr et al reported that airway epithelium exposed to smoke significantly reduced hBD-2 and antimicrobial activity in vitro. In addition, former or current smokers with pneumonia had decreased concentration of hBD-2 in pharyngeal washing fluid compared with non-smokers.43
Our study has several strengths, including analysing ASL samples from the same subjects before and after randomised intervention. We also recognise that we have several limitations such as the small number of samples in the subgroup analysis and the indirect approach to assess the role of LL-37 in the ASL antimicrobial activity using a neutralising antibody. Finally, it is conceivable that smokers require even higher doses of vitamin D3 and/or a longer duration of treatment.
Conclusions
We provide evidence that vitamin D3 supplementation for 90 days increases ASL antimicrobial activity in non-smoking humans. We presume that this effect occurs via an increase in ASL AMPs. Conversely, smoking participants did not improve their ASL antimicrobial activity, and we propose that the mechanism is, in part, due to impaired conversion of vitamin D3 to its active form leading to a decreased expression of airway AMPs. Future studies with a higher number of subjects are required to confirm the effect modification of smoking in the ASL antimicrobial activity in response to vitamin D3 in vivo, as well as the effect of active vitamin D3 in the airway antimicrobial activity of smokers.
Supplementary Material
Supplementary dataAcknowledgments
The authors would like to thank Martha M Monick and Gary H Hunninghake for providing the samples of the Human Lung Responses to Respiratory Pathogens study. We also acknowledge Linda Powers for her technical assistance with the initial processing of ASL samples. In addition, we would like to thank Peter Taft and Julio Noriega for providing editing support.
References
Footnotes
Contributors Conception and design: APC, AKG, LGVB, JZ. Performing the experiments: LGVB, MC, OGVC, AAP. Analysis and interpretation: LGVB, APC, AKG, JZ. Drafting the manuscript for important intellectual content: LGVB, APC, AKG, JZ.
Competing interests None declared.
Patient consent Detail has been removed from this case description/these case descriptions to ensure anonymity. The editors and reviewers have seen the detailed information available and are satisfied that the information backs up the case the authors are making.
Ethics approval IRB# 200607708.
Provenance and peer review Not commissioned; externally peer reviewed.
Data sharing statement Microarray analysis of alveolar macrophages is available to anybody by accessing Gene Expression Omnibus (GEO) accession number: GSE56583. In addition, anybody can contact either corresponding authors, Alicia Gerke at alicia-gerke@uiowa.edu or Alejandro Comellas at alejandro-comellas@uiowa.edu for any enquiries about the manuscript or data reported.